- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
№ 4 Основні принципи класифікації мікробів. 7 страница
| № 59 Імуноглобуліни, структура і функції. Природа імуноглобулінів. У відповідь на введення антигену імунна сістема виробляє антитіла - білки, здатні специфічно со-единящей з антигеном, що викликав їх освіту, і таким чином брати участь в імунологічних реакціях. Ставляться антітела до γ -глобулінів, т. Е. Найменш рухомий в електріческом поле фракції білків сироватки крові. В організмі γ -глобуліни виробляються особливими клітинами - плазмоцитами. γ -глобуліни, несучі функції антитіл, отримали назву іммуноглобулінов і позначаються символом Ig. Отже, антитіла - це імуноглобуліни, що виробляються у відповідь на введення антігена і здатні специфічно взаємодіяти з цим же антигеном. Функції. Первинна функція полягає у взаімодсйствіі їх активних центрів з комплементарними їм детермінантамі антигенів. Вторинна функція полягає в їх здатності: • пов'язувати антиген з метою його нейтралізації та елімінації з організму, т. Е. Брати участь у формуванні защіти від антигену; • брати участь у розпізнаванні «чужого» антигену; • забезпечувати кооперацію імунокомпетентних клітин (макрофагов, Т- і В-лімфоцитів); • брати участь у різних формах імунної відповіді (фагоцітоз, кілерні функція, ГНТ, ГЗТ, імунологічна толерантность, імунологічна пам'ять). Структура антитіл. Білки імуноглобулінів за хімічним складом відносяться до гликопротеидам, так як складаються з протеіна і Сахаров; побудовані з 18 амінокислот. Мають видові відмінності, пов'язані головним чином з набором амінокислот. Їх молекули мають циліндричну форму, їх видно в електронному мікроскопі. До 80% імуноглобулінів мають константу седиментації 7S; стійкі до слабких кіслотам, лугів, нагрівання до 60 ° С. Виділити іммуноглобуліни з сироватки крові можна фізичними і хімічними методамі (електрофорез, Ізоелектрична осадження спиртом і кислотами, висолювання, аффинная хроматографія та ін. ). Ці методи використовують у виробництві при приготуванні імуно-біологічних препаратів. Імуноглобуліни за структурою, антигенними та іммунобіологіческім властивостям поділяються на п'ять класів: IgM, IgG, IgA, IgE, IgD. Імуноглобуліни М, G, А мають подкласси. Наприклад, IgG має чотири підкласу (IgG,, IgG2, IgG3, IgG4). Всі класи і підкласи розрізняються за амінокіслотной послідовності. Молекули імуноглобулінів всіх п'яти класів складаються з поліпептидних ланцюгів: двох однакових важких ланцюгів Н і двох однакових легких ланцюгів - L, з'єднаних між собою дисульфідними містками. Відповідно кожному класу імуноглобулінів, тобто М, G, A, E, D, разлічают п'ять типів важких ланцюгів: μ (мю), γ (гамма), α (альфа), ε (епсилон) і Δ (дельта), що розрізняються за антигенності. Легкі ланцюги всіх п'яти класів є загальними і бувають двох типів: κ (каппа) і λ (лямбда); L-ланцюги імуноглобулінів різних класів можуть вступати в з'єднання (рекомбінуватися) як з гомологічнимі, так і з гетерологічними Н-ланцюгами. Однак в одній і тій же молекулі можуть бути тільки ідентичні L-ланцюга (κ або λ ). Як в Н-, так і в L-ланцюгах є вариабельная - V область, в якій послідовність амінокіслот непостійна, і константная - З область з постійним набором амінокислот. У легких і важких ланцюгах розрізняють NH2- і СООН-кінцеві групи. При обробці γ -глобуліну меркаптоетанолом руйнуються дисульфідні зв'язки і молекула імуноглобуліну розпадається на окремі ланцюги поліпептидів. При впливі протеолітичних ферментом папаїном імуноглобулін розщеплюється на три фрагменти: зо два не кристалізуються, містять детермінантні групи до антигену і названих Fab-фрагментами I і II і один кристалізується Fc-фрагмент. FabI- і FabII-фрагменти подібні за властивостями та амінокислотним складом і відрізняються від Fc-фрагмента; Fab-і Fc-фрагменти є компактними утвореннями, соедіненнимі між собою гнучкими ділянками Н-ланцюги, завдяки чому молекули імуноглобуліну мають гнучку структуру. Як Н-ланцюги, так і L-ланцюга мають окремі, лінійно связанние компактні ділянки, названі доменами; в Н-ланцюги їх по 4, а в L-ланцюга - по 2. Активні центри, або детермінанти, які форміруются в V-областях, займають приблизно 2% поверхні молекули імуноглобуліну. У кожній молекулі є дві детермінанти, що відносяться до гіперваріабельні ділянкам Н-і L-ланцюгів, т. Е. Кожна молекула імуноглобуліну може связать дві молекули антигену. Тому антитіла є двухвалентнимі. Типовою структурою молекули імуноглобуліну є IgG. Інші класи імуноглобулінів відрізняються від IgG додатковими елементами організації їх молекули. У відповідь на введення будь-якого антигену можуть вироблятися антитіла всіх п'яти класів. Зазвичай спочатку виробляється IgM, потім IgG, інші - дещо пізніше. |
№ 60 Класи імуноглобулінів, їх характеристика. Імуноглобуліни за структурою, антигенними та іммунобіологіческім властивостям поділяються на п'ять класів: IgM, IgG, IgA, IgE, IgD. Імуноглобулін класу G. ізотипів G випадках становить основну масу Ig сироватки крові. На його частку припадає 70-80% всіх сивороточ-них Ig, при цьому 50% міститься в тканинної рідини. Середній вміст IgG в сиворотке крові здорової дорослої людини 12 г / л. Період напіврозпаду IgG - 21 день. IgG - мономер, має 2 антигензв'язуючих центру (може одночасно связать 2 молекули антигену, отже, його валентність дорівнює 2), молекулярну масу близько 160 кДа і константу седиментації 7S. Розрізняють підтипи Gl, G2, G3 і G4. Синтезується зрілими В-лімфоцитами і плазматичними клітинами. Добре визна-деляется в сироватці крові на піку первічного і при вторинному імунній відповіді. Має високу аффинностью. IgGl і IgG3 пов'язують комплемент, причому G3 актівнее, ніж Gl. IgG4, подібно IgE, володіє цітофільностью (тропностью, або сродством, до огрядним клітинам і базофілам) і участвует у розвитку алергічної реакції I типу. У імунодіагностичних реакціях IgG може проявляти себе що не-повне антитіло. Легко проходить через плацентарний барьер і забезпечує гуморальний імунітет новонародженого в перші 3-4 місяці життя. Здатний також виділятися в секрет слізістих, у тому числі в молоко шляхом дифузії. IgG забезпечує нейтралізацію, опсонізації та маркування антигену, осуществляет запуск комплемент-опосередкованого цітоліза і антителозависимой клітинно-опосередкованої цитотоксичності. Імуноглобулін класу М. Найбільш крупная молекула з усіх Ig. Це пентамер, кото рий має 10 антигензв'язуючих центрів, т. Е. Його валентність дорівнює 10. Молекулярна маса його близько 900 кДа, константа седіментаціі 19S. Розрізняють підтипи Ml і М2. Важкі ланцюги молекули IgM на відміну від інших ізотипів побудовані з 5 доменів. Період напіврозпаду IgM - 5 днів. На його частку припадає близько 5-10% усіх сироваткових Ig. Середній вміст IgM в сироватці крові здорової дорослої людини становить близько 1 г / л. Цей рівень у людини досягається вже до 2-4-річного віку. IgM філогенетично - найбільш стародавній імуноглобулін. Синтезується предшественнікамі і зрілими В-лімфоцитами. Утворюється на початку первинної імунної відповіді, також першим починає сінтезіроваться в організмі новонародженого - определяется вже на 20-му тижні внутрішньоутробного розвитку. Має високу авідності, найбільш ефективний активатор комплементу по классіческому шляху. Бере участь у формуванні сироваткового і секреторного гуморального імунітету. Будучи полімерної молекулою, що містить J-ланцюг, може утворювати сек-реторную форму і виділятися в секрет слізістих, у тому числі в молоко. Велика частина нормальних антитіл і ізоагглютініни відноситься до IgM. Не проходить через плаценту. Виявлення специфічних антитіл изотипа М в сиворотке крові новонародженого вказує на колишню внутрішньоутробну інфекцію або дефект плаценти. IgM забезпечує нейтралізацію, опсонізації та маркування антигену, осуществляет запуск комплемент-опосередкованого цітоліза і антителозависимой клітинно-опосередкованої цитотоксичності. Імуноглобулін класу А. Існує в сивороточной і секреторною формах. Близько 60% всіх IgA міститься в секретах слизових. Сироватковий IgA: На його частку пріходітся близько 10-15% всіх сироваткових Ig. У сироватці крові здорової дорослої человека міститься близько 2, 5 г / л IgA, максимум досягається до 10-річного віку. Період напіврозпаду IgA - 6 днів. IgA - мономер, має 2 антигензв'язуючих центру (т. Е. 2-валентний), молекулярную масу близько 170 кДа і константу седіментаціі 7S. Розрізняють підтипи А1 і А2. Синтезується зрілими В-лімфоцитами і плазматичними клітинами. Добре определяется в сироватці крові на піку первічного і при вторинному імунній відповіді. Має високу аффинностью. Може бути неповним антитілом. Не пов'язує комплемент. Не проходить через плацентарний бар'єр. IgA забезпечує нейтралізацію, опсоно-зацію та маркування антигену, осуществляет запуск антителозависимой клітинно-опос-редованного цитотоксичности. Секреторний IgA: На відміну від сивороточного, секреторний sIgA існує в полімерній формі у вигляді ди- або тримера (4- або 6-валентний) і містить J- і S-пeптіди. Молекулярна маса 350 кДа і вище, константа седиментації 13S і вище. Синтезується зрілими В-лімфоцитами і їх потомкамі - плазматичними клітинами соответствующей спеціалізації тільки в пределах слизових і виділяється в їхні секрети. Обсяг продукції може досягати 5 г на добу. Пул slgA вважається найчисленнішим в організмі - його кількість перевищує сумарний вміст IgM і IgG. У сироватці крові не виявляється. Секреторна форма IgA - основний фактор специфічного гуморального місцевого імунітету слизових оболонок шлунково-кишкового тракту, сечостатевої системи і респіраторного тракту. Завдяки S-ланцюга він стійкий до дії протеаз. slgA НЕ актівірует комплемент, але ефективно зв'язується з антигенами і нейтралізує їх. Він препятствует адгезії мікробів на епітеліальних клітинах і генералізації інфекції в пределах слизових. Імуноглобулін класу Е. Називають так реагинов. Вміст у сироватці крові вкрай невисоко - приблизно 0, 00025 г / л. Виявлення вимагає застосування спеціальних високочутливих методів діагностікі. Молекулярна маса - близько 190 кДа, константа седиментації - приблизно 8S, мономер. На його частку припадає близько 0, 002% всіх циркулюючих Ig. Цей рівень достігается до 10-15 рокам життя. Синтезується зрілими В-лімфоцитами і плазматичними клітинами преімущественно в лімфоїдної тканини бронхолегеневого дерева і ШКТ. Не пов'язує комплемент. Не проходить через плацентарний бар'єр. Володіє вираженной цітофільностью - тропність до огрядним клітинам і базофілам. Бере участь у розвитку гіперчутливості негайного типу - реакція I типу. Імуноглобулін класу D. Відомостей про Ig даного изотипа не так багато. Практично повністю утримується в сироватці крові в концентрації близько 0, 03 г / л (близько 0, 2% від загального числа циркулюючих Ig). IgD має молекулярну масу 160 кДа і константу седіментаціі 7S, мономер. Не пов'язує комплемент. Не проходить через плацентарний бар'єр. Є рецептором попередників В-лімфоцитів. |
№ 61 Антигени: визначення, основні властивості. Антігени бактеріальної клітини. Антиген - це біополімер органічної природи, генетично чужорідний для макроорганізму, який при потраплянні в останній розпізнається його імунною системою і викликає імунні реакції, спрямовані на його усунення. Антигени мають ряд характерних властивостей: антигенностью, специфічністю і иммуногенностью. Антигенность. Під антигенностью розуміють потенціальную здатність молекули антигену актівіровать компоненти імунної системи і специфічно взаємодіяти з факторамі імунітету (антитіла, клон ефекторних лімфоцитів). Іншими словами, антиген дол жен виступати специфічним подразником по відношенню до імунокомпетентним клеткам. При цьому взаємодія компоненти імунної системи відбувається не з усією молекулою одночасно, а тільки з її небольшім ділянкою, який отримав назву «антигенная детермінанта», або «епітоп». Чужорідність є обов'язковим условіем для реалізації антигенності. За цим критерієм система набутого імунітету диференціює потенційно небезпечні об'екти біологічного світу, синтезовані з чужорідної генетичної матриці. Поняття «чужорідність» відносне, так як имму-нокомпетентние клітини не здатні напрямую аналізувати чужорідний генетичний код. Вони сприймають лише опосередковану інформацію, яка, як у дзеркалі, відображена в молекулярній структурі речовини. Імуногенність - потенційна способность антигену викликати по відношенню до себе в макроорганизме специфічну защітную реакцію. Ступінь імуногенності залежить від ряду факторів, які можна об'єднати в три групи: 1. Молекулярні особливості антигену; 2. Кліренс антигену в організмі; 3. Реактивність макроорганізму. До першої групи факторів віднесені природа, хімічний склад, молекулярна вага, структура і деякі інші характеристики. Імуногенність в значній мірі завісіт від природи антигену. Важлива також оптична ізомерія амінокислот, складових молекулу білка. Велике значення має розмір і молекулярна маса антигену. На ступінь імуногенності також оказивает вплив просторова структура антігена. Виявилася також суттєвої стерическое стабільність молекули антигену. Ще однією важливою умовою імуно-генності є розчинність антигену. Друга група факторів пов'язана з динамікою надходження антигену в організм і його виведенія. Так, добре відома залежність імуногенності антигену від способу його введення. На імунну відповідь впливає кількість поступающего антигену: чим його більше, тим більше виражений імунну відповідь. Третя група об'єднує чинники, определяющіе залежність імуногенності від состоянія макроорганізму. У цьому зв'язку на первий план виступають спадкові чинники. Специфічністю називають здатність антігена індукувати імунну відповідь до строго певного Епітопи. Ця властивість обусловлено особливостями формування іммунного відповіді - необхідна компліментарність рецепторного апарату імунокомпетентних клітин до конкретної антигенної детермінанті. Тому специфічність антигену в чому визначається властивостями складових його епітопів. Однак при цьому слід враховувати умовність кордонів епітопів, їх структурний різноманітність і гетерогенність клонів антігенреактівние лімфоцітовой специфічності. У результаті цього організм на антигенное раз Роздратування завжди відповідає поліклональними іммунним відповіддю. Антігени бактеріальної клітини. У структурі бактеріальної клітини разлічают жгутиковие, соматичні, капсульні і деякі інші антигени. Жгутикові, або Н-антигени, локалізуются в локомоторними апараті бактерій - їх джгутиками. Вони являють собою епітопи скорочувального білка флажеліну. При нагреваніі флажеліну денатурує, і Н-антиген втрачає свою специфічність. Фенол не діє на цей антиген. Соматический, або О-антиген, пов'язаний з клітинною стінкою бактерій. Його основу составляют ЛПС. О-антиген проявляє термостабільние властивості - він не руйнується при тривалому кип'ятінні. Однак соматіческій антиген піддається дії альдегідів (наприклад, формаліну) і спиртів, які порушують його структуру. Капсул'ние, або К-антигени, розташовуються на поверхні клітинної стінки. Зустрічаються у бактерій, що утворюють капсулу. Як правило, К-антигени складаються з кислих полісахаридів (уроновие кислоти). У той же час у бацили сибірської виразки цей антиген побудований з по-ліпептідних ланцюгів. За чутливості до нагрівання розрізняють три типу К-антигену: А, В, і L. Найбільша термостабільність характерна для типу А, він не денатурує навіть при тривалому кип'ятінні. Тип В видержівает нетривалий нагрівання (близько 1 години) до 60 " С. Тип L швидко руйнується при цій температурі. Тому часткове удаленіе К-антигену можливо шляхом тривалого кип'ятіння бактеріальної культури. На поверхні збудника черевного тіфа та інших ентеробактерій, які обладают високою вірулентністю, можна обнаружіть особливий варіант капсульного антигену. Він отримав назву антигену вірулентності, або Vi-антигену. Виявлення цього антігена або специфічних до нього антитіл імеет велике діагностичне значення. Антигенними властивостями володіють також бактеріальні білкові токсини, ферменти і деякі інші білки, які секретуються бактеріями в навколишнє середовище ( приклад, туберкулін). При взаємодії зі специфічними антитілами токсини, ферменти та інші біологічно активні молекули бактеріального походження втрачають свою активність. Правцевий, діфтерійний і ботулінічний токсини відносяться до числа сильних повноцінних антигенів, поетому їх використовують для отримання анатоксінов для вакцинації людей. В антигенному складі деяких бактерій виділяється група антигенів з сильно вираженной иммуногенностью, чия біологічна активність відіграє ключову роль в формірованіі патогенності збудника. Зв'язування таких антигенів специфічними антитілами практично повністю інактивує вірулентние властивості мікроорганізму і обеспечівает імунітет до нього. Описувані антігени отримали назву протективних. Вперше протективний антиген був виявлений в гнійному виділеннях карбункула, викликаного баціллой сибірської виразки. Ця речовина является субодиницею білкового токсину, яка відповідальна за активацію інших, власне вірулентних субодиниць - так званого набрякового і летального факторів. | |
№ 62 Антитілоутворення: первинний і вторинний ответ.
Здатність до утворення антітел з'являється у внутрішньоутробному періоді у 20-тижневого ембріона; після народження починається власна продукція імуноглобулінів, яка збільшується до настання зрелого віку та дещо знижується до старості. Динаміка образованія антитіл має різний характер залежно від сили антигенного впливу (дози антигену), частоти впливу антигену, стану організму і його імунної системи. При первинному і повторному введенні антигену динаміка антитілоутворення також різна і протікає в кілька стадій. Виділяють латентну, логарифмічну, стаціонарную фазу і фазу зниження. 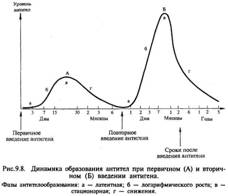 В латентній фазі відбуваються переробка та подання антигену імунокомпетентним клітинам, розмноження клону клітин, спеціалізованого на вироблення антитіл до даного антигену, починається синтез антітел. У цей період антитіла в крові не виявляються.
Під час логарифмічною фази синтезовані антитіла висво-няється з плазмоцитів і надходять в лімфу та кров.
У стаціонарной фазі кількість антитіл досягає максимуму і ста-білізіруется, потім настає фаза зниження рівня антитіл. При первинному введенні антигену (первинну імунну ответ) латентна фаза становить 3-5 діб, логарифмічна - 7 15 сут, стаціонарна - 15-30 сут і фаза зниження - 1-6 місяців і більше. Особливістю первинного імунної відповіді є те, що спочатку синтезується IgM, а потім IgG.
На відміну від первинної імунної відповіді при вторинному введенні антигену (вторинний імунну відповідь) латентний період укорочений до декількох годин або 1-2 діб, логаріфміческая фаза характеризується швидким наростанням і значітельно більш високим рівнем антитіл, який у наступних фазах довго утримується і повільно, іноді протягом несколькіх років, знижується. При вторинному імунній відповіді на відміну від первинного синтезуються головним чином IgG.
Така відмінність динаміки антитілоутворення при первічном і вторинному імунній відповіді пояснюється тим, що після первинного введення антигену в імунній системі форміруется клон лімфоцитів, які несуть імунологічну пам'ять про даний антигене. Після повторної зустрічі з цим же антігеном клон лімфоцитів з імунологічної пам'яттю швидко размножается і інтенсивно включає процес антителогенеза.
Дуже швидке й енергійне антителообразование при повторной зустрічі з антигеном використовується в практичних цілях при необхідності отримання високих титрів антитіл при проізводстве діагностичних і лікувальних сироваток від іммунізірованних тварин, а також для екстреного створення іммунітета при вакцинації.
В латентній фазі відбуваються переробка та подання антигену імунокомпетентним клітинам, розмноження клону клітин, спеціалізованого на вироблення антитіл до даного антигену, починається синтез антітел. У цей період антитіла в крові не виявляються.
Під час логарифмічною фази синтезовані антитіла висво-няється з плазмоцитів і надходять в лімфу та кров.
У стаціонарной фазі кількість антитіл досягає максимуму і ста-білізіруется, потім настає фаза зниження рівня антитіл. При первинному введенні антигену (первинну імунну ответ) латентна фаза становить 3-5 діб, логарифмічна - 7 15 сут, стаціонарна - 15-30 сут і фаза зниження - 1-6 місяців і більше. Особливістю первинного імунної відповіді є те, що спочатку синтезується IgM, а потім IgG.
На відміну від первинної імунної відповіді при вторинному введенні антигену (вторинний імунну відповідь) латентний період укорочений до декількох годин або 1-2 діб, логаріфміческая фаза характеризується швидким наростанням і значітельно більш високим рівнем антитіл, який у наступних фазах довго утримується і повільно, іноді протягом несколькіх років, знижується. При вторинному імунній відповіді на відміну від первинного синтезуються головним чином IgG.
Така відмінність динаміки антитілоутворення при первічном і вторинному імунній відповіді пояснюється тим, що після первинного введення антигену в імунній системі форміруется клон лімфоцитів, які несуть імунологічну пам'ять про даний антигене. Після повторної зустрічі з цим же антігеном клон лімфоцитів з імунологічної пам'яттю швидко размножается і інтенсивно включає процес антителогенеза.
Дуже швидке й енергійне антителообразование при повторной зустрічі з антигеном використовується в практичних цілях при необхідності отримання високих титрів антитіл при проізводстве діагностичних і лікувальних сироваток від іммунізірованних тварин, а також для екстреного створення іммунітета при вакцинації.
|
№ 63 Иммунологическая пам'ять. Імунологічна толерантность. Імунологічна пам'ять. При повторній зустрічі з антигеном орга-нізм формує більш активну і швидку імунну реакцію - вторинний імунну відповідь. Цей феномен отримав назву имму-технологічною пам'яті. Імунологічна пам'ять має високую специфічність до конкретного антігену, поширюється як на гуморальну, так і клітинну ланку імунітету і обусловлена В- і Т-лімфоцитами. Вона образуется практично завжди і зберігається роками і навіть десятиліттями. Завдяки ній наш організм надійно захищений від повторних антигенних інтервенцій. На сьогоднішній день розглядають два найбільш вірогідних механізму формірованія імунологічної пам'яті. Один з них передбачає тривале збереження антігена в організмі. Цьому є безліч прикладів: інкапсульований збудник туберкульозу, персистирующие віруси кору, поліомієліту, вітряної віспи та деякі інші патогени тривалий час, іноді все життя, зберігаються в організмі, поддержівая в напрузі імунну систему. Ймовірно також наявність довгоживучих дендрітних АПК, здатних довго сохранять і презентировать антиген. Інший механізм передбачає, що в процессе розвитку в організмі продуктивного іммунного відповіді частину антігенореактівних Т- або В-лімфоцитів диференціюється в малі покоящіеся клітини, або клітини імунологічної пам'яті. Ці клітини відрізняються високою спе-ціфічностью до конкретної антигенної детермінанте і великою тривалістю життя (до 10 років і більше). Вони активно рециркулируют в організмі, розподіляючись в тканинах і органах, але постійно повертаються в місця свого проісхожденія за рахунок хомінгових рецепторів. Це забезпечує постійну готовність імунної системи реагувати на повторний контакт з антигеном по вторинному типу. Феномен імунологічної пам'яті широко використовується в практиці вакцинації людей для створення напруженого імунітету і під-тримання його тривалий час на захисному рівні. Здійснюють це 2-3-кратними прівівкамі при первинної вакцинації і періоди-дических повторними введеннями вакцінного препарату - ревакцинациями. Однак феномен імунологічної пам'яті має і негативні сторони. Наприклад, повторна спроба трансплантувати вже одного разу відторгнуту тканину викликає биструю і бурхливу реакцію - криз відторгнення. Імунологічна толерантность - явленіе, протилежне імунної відповіді та імунологічної пам'яті. Проявляється вона відсутністю специфічного продуктивного імунної відповіді організму на антиген у зв'язку з нездатністю його розпізнавання. На відміну від імуносупресії іммунологіческая толерантність передбачає початкову ареактівность імунокомпетентних клітин до певного антигену. Імунологічну толерантність визивают антигени, які отримали назву толерогена. Ними можуть бути практично всі речовини, однак найбільшою толерогенниє володіють полісахариди. Імунологічна толерантність бивает вродженої та набутої. Прикладом вродженої толерантності є отсутствіе реакції імунної системи на свої власні антигени. Придбану толерантность можна створити, вводячи в організм речовини, що пригнічують імунітет (імунодепресанти), або ж шляхом введення антигену в ембріональному періоді або в перші дні після народження індивідуума. Придбана толерантність може бути активною і пассівной. Активна толерантність створюється путем введення в організм толерогена, який формує специфічну толерантність. Пасивну толерантність можна викликати веществамі, що гальмують біосинтетичну або проліферативну активність імунокомпетентних клітин (антилімфоцитарну сироватка, цитостатики та ін. ). Імунологічна толерантність отлічается специфічністю - вона спрямована до строго певних антигенів. За ступенем рас-пространение розрізняють поливалентную і розщеплену толерантність. Полівалентна толерантність виникає одночасно на всі антигенні детермінанти, що входять до состав конкретного антигену. Для розщепленої, або моновалентної, толерантності характерна виборча несприйнятливість якихось окремих антигенних детермінант. Ступінь прояву імунологічної толерантності істотно залежить від ряду властивостей макроорганізму і толерогена. Важливе значення в індукції іммунологіческой толерантності мають доза антігена і тривалість його впливу. Розрізняють високодозову і низькодозового толерантність. Високодозову толерантність викликають введенням великих кількостей ви-сококонцентрірованного антигену. Низькодозового толерантність, навпаки, викликається дуже малою кількістю ви-сокогомогенного молекулярного антигену. Механізми толерантності різноманітні і до кінця не розшифровані. Відомо, що її основу складають нормальні процеси регуляції імунної системи. Виділяють три найбільш ймовірні причини розвитку іммунологіческой толерантності: 1. Елімінація з організму антигенспецифических клонів лімфоцитів. 2. Блокада біологічної активності їм-мунокомпетентних клітин. 3. Швидка нейтралізація антигену антітеламі. Феномен імунологічної толерантності має велике практичне значення. Він використовується для вирішення багатьох важливих проблем медицини, таких як пересадка органов і тканин, придушення аутоімунних реакцій, лікування алергій та інших патологіческіх станів, пов'язаних з агрессівним поведінкою імунної системи. |
| |||||||||||||||||||||||||||||||||||||||||||
| № 65 Механізми гіперчутливості уповільненого типу. Клініко-діагностичне значення. До алергічних реакціям відносять два типи реагування на чужорідну речовину: гіперчутливість негайного типу (ГНТ) і гіперчутливість сповільненого типу (ГСТ). До ГНТ відносяться алергічні реакції, які проявляються вже через 20-30 хв після повторної зустрічі з антигеном, а до ГЗТ - реакції, що виникають не раніше ніж через 24-48 ч. Механізм і клініческіе прояви ГНТ і ГЗТ різні. ГНТ пов'язана з виработкой антитіл, а ГЗТ - з клітинними реакціями. ГЗТ вперше описана Р. Кохом (1890). Ця форма прояву не пов'язана з антитілами, опосередкована клітинними механізмамі за участю Т-лімфоцитів. До ГЗТ відносяться наступні форми прояви: туберкулінова реакція, уповільнена алергія до білків, контактна алергія. На відміну від реакцій I, II і III типів реакції IV типу не связани з антитілами, а обумовлені клітинними реакціями, насамперед Т-лімфоцитами. Реакції сповільненого типу можуть виникати при сенсибілізації організму: 1. Мікроорганізмами і мікробними антигенами (бактеріальнимі, грибковими, протозойними, вірусними); 2. гельмінтів; 3. Природними і штучно синтезованими гаптенами (лікарські препарати, барвники); 4. Деякими білками. Отже, реакція уповільненого типу може визиваться практично всіма антигенами. Але найбільш яскраво вона проявляется на введення полісахаридів, низькомолекулярних пептідов, т. Е. Малоіммуногенних антигенів. При цьому реакцію викликають малі дози антигенів і найкраще при внутрішньошкірне введення. Механізм алергічної реакції цього типу полягає в сен-сібілізаціі Т-лімфоцитів-хелперів антигеном. Сенсибілізація лімфоцитів викликає виділення медіаторів, зокрема інтерлейкіну-2, які активують макрофаги і тим самим вовлекают в процес руйнації антигену, що викликав сенсибілізацію лімфоцитів. Цитотоксичність виявляють також і самі Т-лімфоцити. Про роль лімфоцитів у виникненні аллергій клітинного типу свідчать можливість передачі аллергіі від сенсибілізованої тварини несенсибілізованих за допомогою введення лімфоцитів, а також придушення реакції за допомогою антилімфоцитарну сироватки. Морфологічна картина при алергіях клітинного типу носіт запальний характер, обумовлений реакцією лімфоцітов і макрофагів на утворений комплекс антигену з сен-сібілізірованнимі лімфоцитами. Алергічні реакції клітинного типу проявляються у вигляді туберкулінової реакції, сповільненої алергії до білків, контактной алергії. Туберкулінова реакція виникає через 5-6 год після внутрішньошкірного введення сенсибілізованим туберкульозної палочкой тваринам або людині туберкуліну, тобто. Е. Антигенів туберкулезной палички. Виражається реакція у вигляді почервоніння, пріпухлості, ущільнення на місці введення туберкуліну. Сопровождается іноді підвищенням температури тіла, лимфопенией. Развітіе реакції досягає максимуму через 24-48 ч. Туберкуліновая реакція використовується з діагностичною метою для виявленія захворювань на туберкульоз або контактів організму з туберкульозною паличкою. Уповільнена алергія виникає при сенсибілізації малимі дозами білкових антигенів з ад'ювантом, а також кон'-гатамі білків з гаптенами. У цих випадках алергічна реакція виникає не раніше ніж через 5 днів і триває 2-3 тижнів. Мабуть, тут грають роль уповільнене дію кон'югованих білків на лімфоїдну тканину і сенсибілізація Т-лімфо-цитов. Контактна алергія виникає, якщо антигенами є низькомолекулярні органічні та неорганічні речовини, які в організмі з'єднуються з білками, утворюючи кон'юга-ти. Кон'юговані з'єднання, виконуючи роль гаптенов, визивают сенсибілізацію. Контактна алергія може виникати при тривалому контакті з хімічними речовинами, в тому числі фармацевтичними препаратами, фарбами, косметіческімі препаратами (губна помада, фарба для вій). Проявляется контактна алергія у вигляді всіляких дерматитів, т. Е. Поразок поверхневих шарів шкіри. Значення. Всі реакції гіперчутливості, у тому числі і ГЗТ мають велике значення. Їх механізми лежать в основі запалення, яке сприяє локалізації інфекційного агента чи іншого антигену в межах певних тканин і формуванню повноцінної імунної реакції захисного характеру. |
№ 66 Алергічні проби, їх сутності, застосування. Алергічні проби - біологічні реакції для діагностики ряду захворювань, засновані на підвищеній чутливості організму, викликаної алергеном. При багатьох інфекційних захворюваннях за рахунок активації кле-точного імунітету розвивається підвищена чутливість організму до збудників і продуктів їх життєдіяльності. На цьому засновані алергічні проби, використовувані для діагностікі бактеріальних, вірусних, протозойних інфекцій, мікозов і гельмінтозів. Алергічні проби володіють спеціфічностью, але нерідко вони бувають позитивними у перехворілих і щеплених. Всі алергічні проби поділяють на дві групи - проби in vivo і in vitro. До першої групи {in vivo) відносяться шкірні проби, осуществляемие безпосередньо на пацієнті і виявляють алергію негайного (через 20 хв) та уповільненого (через 24 - 48 год) типів. Алергічні проби in vitro засновані на виявленні сенсібілізаціі поза організмом хворого. Їх застосовують тоді, коли з тих чи інших причин не можна провести шкірні проби, або в тих випадках, коли шкірні реакції дають неясні результати. Для проведення алергічних проб використовують алергени - діагностичні препарати, призначені для виявлення специфічної сенсибілізації організму. Інфекційні аллергени, використовувані в діагностиці інфекційних заболеваній, являють собою очищені фільтрати бульйонних культур, рідше суспензії убитих мікроорганізмів або АГ, виділені з них. Шкірні проби. Інфекційні алергени вводять, як правило, внутрішньошкірно або нашкірному, шляхом втирання в скаріфіціровать ділянки шкіри. При внутрикожном способі в середню третину передньої поверхності передпліччя спеціальною тонкою голкою вводять 0, 1 мл аллергена. Через 28 - 48 год оцінюють результати реакції ГЗТ, визначаючи на місці введення розміри папули. Неінфекційні алергени (пилок рослин, побутовий пил, харчові продукти, лікарські та хімічні препарати) вводять в шкіру уколом (прик-тест), накожно шляхом скарификации і втирання або внутрішньошкірної ін'єкцією розведеного розчину алергену. В якості негативного контролю використовують їхн, в якості позитивного - розчин гістаміну. Результати учітивают протягом 20 хв (ГНТ) за величиною папули (іноді до 20 мм в діаметрі), наявності набряку та свербежу. Внутрішньошкірні проби ставлять у випадку негативного чи сумнівного результату прик-тесту. У порівнянні з останнім, дозу алергену зменшують в 100-5000 разів. Шкірні проби на наявність ГЗТ широко застосовують для виявленія інфікованості людей мікобактеріями туберкульозу (проба Манту), збудниками бруцельозу (проба Бюрне), лепри (реакція Мітсуди), туляремії, сапа, актиномикоза, дерматомікозів, токсоплазмозу, деяких гельмінтозів та ін. Проби in vitro. Ці методи дослідження безпечні для хворого, достаточно чутливі, дозволяють кількісно оцінити рівень алергізації організму. В даний час розроблені тести для визначення сенсібілізаціі, засновані на реакціях Т- і B-лімфоцитів, тканевих базофілів, виявленні загальних специфічних IgE в сиворотке крові та ін. До них відносяться реакції гальмування міграції лейкоцитів і бласттрансформации лімфоцитів, специфічне розеткоутворення, базофільний тест Шеллі, реакція дегрануляції тканинних базофілів, а також аллергосорбентние методи (визначення специфічних IgE в сироватці крові). Реакція гальмування міграції лейкоцитів (РТМЛ). РТМЛ основана на придушенні міграції моноцитів та інших лейкоцитів під дією медіаторів, що виробляються сенсібілізірованнимі лімфоцитами, у присутності специфічного алергену. Реакція бласттрансформації лімфоцитів (РБТ). В основі цієї реакції лежить здатність нормальних лімфоцитів періферіческой крові вступати в мітоз і перетворюватися на бластні форми при культивуванні їх in vitro під дією специфічних факторов - алергенів і неспецифічних стимуляторів мітогенез - митогенов (фитогемагглютинин, конканавалін А, ліпополісахариди та інші речовини). Реакція специфічного розеткоутворення. Розетки - харак-терни утворення, що виникають in vitro в результаті пріліпанія еритроцитів до поверхні імунокомпетентних клітин. Розеткоутворення може відбуватися спонтанно, оскільки Т-лімфоцити людини містять рецептори до еритроцитів барана. Спон-танное розеткоутворення здорових людей становить 52 - 53% і служить показником функціонального стану Т-лімфоцитів. Цей феномен відтворюється також і в тому випадку, якщо використовують еритроцити, на яких фіксовані соответствующіе алергени. Реакція дегрануляції тканинних базофілів. Методика основана на тому, що під дією алергену відбувається дегрануляція тканинних базофілів щури, попередньо сенсібілізірованних цітофільнимі AT з сироватки крові хворого. Базофільний тест Шеллі. Відомо, що базофільні гранулоцити людини або кролика також дегранулюють в прісутствіі сироватки хворого і алергену, до якого чутливий даний пацієнт. Визначення антитіл класу IgE in vitro. Лабораторна діагностіка захворювань, в основі яких лежить ГНТ, заснована на визначенні аллергенспецифических IgEанті-IgE. При іспользованіі радіоактивної мітки метод носить названіе радіоаллергосорбентний тесту (PACT), але частіше в качестве мітки використовують фермент або флюоресцирующєє речовина (ФАСТ). Час аналізу - 6 - 7 годин. Принцип методу: фіксірованний на твердій основі відомий алерген інкубують з сивороткой крові хворого; знаходяться в сироватці спеціфіческіе IgEанті-IgE зв'язуються з алергеном і, таким чином, залишаються фіксованими на основі і можуть вступати в спеціфіческое взаємодію з додаються міченими анти-IgE. |
№ 67 Гіперчувствітельност' негайного типу. Механізми виникнення, клінічна значимість. Гіперчутливість негайного типу (ГНТ) - ги-перчувствітельность, обумовлена антитілами (IgE, IgG, IgM) проти алергенів. Розвивається через сколько хвилин або годин після дії алергену: расшіряются судини, підвищується їх проникність, развіваются свербіж, бронхоспазм, висип, набряки. Пізня фаза ГНТ дополняется дією продуктів еозинофілів і нейтрофілів. До ГНТ відносяться I, II і III типи алергічних реакцій (по Джеллу і Кумбсу): I тип - анафілактичний, обусловленний головним чином дією IgE; II тип - цитотоксичний, обумовлений дією IgG, IgM; III тип - імунокомплексний, що розвивається при утворенні імунного комплексу IgG, IgM з антигенами. В окремий тип виділяють антірецепторние реакції. Основні типи реакцій гіперчутливості I тип - анафілактичний. При первинному контакті з антігеном утворюються IgE, які прикріплюються Fc-фрагментом до огрядним клітинам і базофілам. Повторно введенний антиген перехресно зв'язується з IgE на клітинах, викликаючи їх дегрануляцію, викид гістаміну та інших медіаторов алергії. Первинне надходження алергену викликає продукцію плазмацітамі IgE, IgG4. Синтезовані IgE прикріплюються Fc-фрагментом до Fc-pe цепторов (FceRl) базофілів в крові і огрядних клітин в слизових оболонках, сполучної тканини. При повторному поступленіі алергену на тучних клітинах і базофілів образуюто комплекси IgE з алергеном (перехресна зшивання FceRl антігеном), що викликають дегрануляцію клітин. Клінічні прояви гіперчутливості I типу. Клінічні прояви гіперчутливості I типу можуть протікати на тлі атопії. Атопія - спадкова схильність до розвитку ГНТ, обумовлена повишенной виробленням IgE-антитіл до алергену, підвищеною кількістю Fc-рецепторів для цих антитіл на огрядних клетках, особливостями розподілу огрядних клітин і повишенной проникністю тканинних бар'єрів. Анафілактичний шок - протікає гостро з розвитком колапсу, набряків, спазму гладкої мускулатури; часто заканчівается смертю. Кропив'янка - збільшується проніцаемость судин, шкіра червоніє, з'являються пухирі, свербіж. Бронхіальна астма - розвиваються запалення, бронхо-спазм, посилюється секреція слизу в бронхах. II тип - цитотоксичний. Антиген, розташований на клітці «впізнається» антитілами класів IgG, IgM. При взаімодействіі типу «клітина-антиген-антитіло» відбувається актівація комплементу і руйнування клітини за трьома напрямками: комплементзавісімий цитолиз; фагоцитоз; антитілозалежна клітинна цитотоксичність. Час реакції - хвилини або години. До II типу гіперчутливості близькі антірецепторние реакції (так званий IV тип гіперчутливості), основою яких є антірецепторние антитіла, приклад антитіла проти рецепторів до гормонів. Клінічні прояви II типу. За II типу гіперчувствітельності розвиваються деякі аутоімунні хвороби, обусловленние появою аутоантитіл до антигенів власних тканин: злоякісна міастенія, аутоіммунная гемолітична анемія, вульгарна пухирчатка, синдром Гудпасчера, аутоімунний гіпертиреоїдизм, інсулінозаві-сімий діабет II типу. Аутоіммунну гемолітичну анемію викликають антітела проти Rh-антигену еритроцитів; еритроцити разрушаются в результаті активації комплементу і фагоцитозу. Лекарственно-індуковані гемолітична анемія, гранулоцитопенія і тромбоцитопенія супроводжуються появленіем антитіл проти ліки - гаптена і цитолизом клеток, що містять цей антиген. III тип - імунокомплексний. Антитіла класів IgG, IgM утворюють з розчинними антигенами імунні комплекси, які активують комплемент. При надлишку антигенів або недоліку комплементу імунні комплекси откладиваются на стінці судин, базальних мембранах, т. Е. Структурах, що мають Fc-рецептори. Первинними компонентами III типу гіперчутливості є розчинні імунні комплекси антиген-антітело і комплемент (анафілатоксин С4а, СЗа, С5а). При ізбитке антигенів або недоліку комплементу імунні комплекси відкладаються на стінках судин, базальних мембранах, тобто структурах, що мають Fc-рецептори. Поврежденія обумовлені тромбоцитами, нейтрофілами, імунними комплексами, комплементом. Залучаються прозапальні цитокіни, включаючи TNF-a та хемокіни. На пізніх стадіях в процес залучаються макрофаги. Реакція може бути загальною (наприклад, сироваткова болезнь) або залучати окремі органи, тканини, включаючи кожу (наприклад, системний еритематозний вовчак, реакція Артюса), нирки (наприклад, вовчаковий нефрит), легкі (наприклад, аспергільоз) або інші органи. Ця реакція може бути обумовлена багатьма мікроорганізмами. Вона розвивається через 3-10 годин після експозиції антигену, як у реакції Артюса. Антиген може бути екзогенні (хроніческіе бактеріальні, вірусні, грибкові або прото-зойние інфекції) або ендогенний, як при системній Ері-тематозной вовчак. Клінічні прояви III типу. Сироваткова хвороба відбувається при введенні високих доз антигену, наприклад кінської протиправцевої сивороткі. Через 6-7 днів у крові з'являються антитіла протів кінського білка, які, взаємодіючи з даним антигеном, утворюють імунні комплекси, откладивающіеся в стінках кровоносних судин і тканинах. Розвиваються сістемние васкуліти, артрити (відкладення комплексів в суставах), нефрит (відкладення комплексів в нирках). Реакція Артюса розвивається при повторному внутрішньошкірне введення антигену, який локально утворює іммунние комплекси з раніше накопиченими антитілами. Проявляется набряком, геморагічним запаленням і некрозом. | |
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|