- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 6 страница
Акустико-латеральная область составляет значительную часть дорсолатеральных отделов продолговатого мозга. Она рассматривается в качестве деривата соматической сенсорной: колонны на основании как своего положения в мозге, так и происхождения рецепторов данной системы из эктодермальных. производных - плакод. Данная область мозга является зоной представительства октаво-латеральной системы, объединяющей органы специализированной механорецепции и их центральные проекции. Октаво-латеральная система на периферии включает в себя сообщающиеся с внешней средой органы боковой линии и изолированный от внешней среды отдел-лабиринт. Первые обнаруживаются только у водных позвоночных и обеспечивают поступление информации о перемещениях как самого животного, так и водной среды. У некоторых групп низших позвоночных имеются специализированные органы боковой линии, чувствительные к электрическим воздействиям (миноги, некоторые рыбы, хвостатые амфибии). Электрорецепторы таких органов адаптированы к восприятию градиента электрического поля и таким образом опосредуют реакции животного как на электрические поля других особей, так и на изменения собственного электрического поля. Октавная подсистема получает
информацию от механорецепторов, лежащих в лабиринте. Лабиринт содержит два функционально различных отдела - вестибулярный аппарат, который обеспечивает восприятие перемещений тела и является органом гравитации, и внутреннее ухо - звуковоспринимающий отдел (последний как самостоятельный отдел у низших позвоночных отсутствует, а его функции выполняют элементы вестибулярного аппарата).
Октавная подсистема обнаруживается у всех позвоночных, органы боковой линии - у всех анамний в личиночной стадии и у большинства взрослых анамний. Менее широко представлена электрорецептивная часть: она отсутствует у голостей, большинства телеостей и некоторых амфибий.
Сходство в структуре и характере развития рецепторов разных отделов октаво-латеральной системы привело к формированию представления об общности их происхождения. Ранее полагали, что механорецепторы канальных органов боковой линии дают начало электрорецепторам и рецепторам внутреннего уха, причем среди последних раньше возникают вестибулярные рецепторы. Слуховая рецепция появляется лишь у переходных к амфибиям форм с образованием передающих аппаратов среднего уха и рецепторов, способных отвечать на звуки, распространяющиеся по воздуху. Переход от водной к наземной среде обитания у многих амфибий знаменуется утратой органов боковой линии, а проекции новой системы адресуются той же области продолговатого мозга, изменяя ее функции. Однако, современные морфологические и физиологические данные свидетельствуют о самостоятельности и параллелизме в развитии каждого компонента октаво-латеральной системы. Иными словами, каждый из них, появившись рано и одновременно в эволюции позвоночных, является "базовым" отделом продолговатого мозга. Изменения, которые затем происходят в каждой из субсистем, включают адаптивные преобразования и центрального представительства (в том числе редукцию центральных отделов системы органов боковой линии с исчезновением соответствующих рецепторов).
Каждая часть октаво-латеральной системы имеет собственные пути проведения, представленные краниальными нервами, а также специализированную область в пределах акустико-латеральной зоны продолговатого мозга.
У животных, обладающих органами боковой линии, в организации путей проведения обнаруживается ряд общих черт. Так, у круглоротых и рыб информация от рецепторного уровня поступает в мозг по трем парам нервов: дорсальному и вентральному передним нервам боковой линии, nn. lineae lateralis anterior dorsalis et ventralis, и заднему нерву, n. lineae lateralis posterior. Чаще всего их рассматривают как специализированные отделы лицевого (передний нерв) и блуждающего (задний нерв) черепномозговых нервов; однако некоторые авторы отстаивают их самостоятельное происхождение.
Центры представительства органов боковой линии расположены в дорсальной и промежуточной зонах акустико-латеральной области и покрыты снаружи весьма характерным образованием - мозжечковым гребнем, crista cerebelli, который является продолжением молекулярного слоя мозжечка (рис. 34, 35).
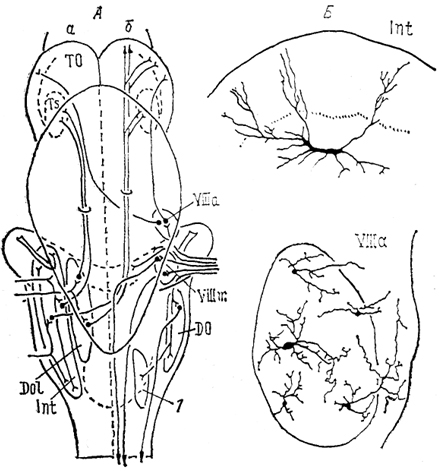
Рис. 36. Схема связей (А) и нейронный состав (Б) ядер октаво-латеральной области хрящевых рыб (Smeets e. а., 1983).
а, б - связи ядер органов боковой линии (а), VIII нерва (б). 1 - ядро бокового канатика.
В пределах мозжечкового гребня формируются контакты нейронов центров боковой линии с параллельными волокнами и аксонами звездчатых клеток мозжечка.
Система связей центров боковой линии изучена недостаточно полно. Показано, что крупные клетки, входящие в их состав и получающие первичные афференты, в свою очередь проецируются на элементы ретикулярной формации. Восходящие проекции в средний мозг обнаружены у пластиножаберных и некоторых телеостей (рис. 36, А).
Степень и характер развития области представительства органов боковой линии имеет ряд отличий у позвоночных разных групп. Так, у миног и пластиножаберных, а также у некоторых костных рыб (хондростей, двоякодышащих и многоперовых) в ее пределах выделяют дорсальное и медиальное ядра, nucll. dorsalis et medialis, иначе переднюю и заднюю доли боковой линии, lobi lineae lateralis anterior et posterior (рис. 29, 34). Полагают, что у пластиножаберных имеется вход В торус среднего мозга через примитивную систему латеральной петли. Однако, по мнению других исследователей, эфференты
дорсального и медиального ядер направляются контрлатерально в субтектальные области.
У миксин, голостей и телеостей проекции органов боковой линии адресованы медиальному ядру акустико-латеральной зоны. У телеостей оно состоит из дорсальной и медиальной частей. Первая более дифференцирована, дендриты ее нейронов проникают в мозжечковый гребень и получают входы от заднего нерва боковой линии (от рецепторов туловищного отдела). Медиальная часть не имеет четких границ и связана с представительством рецепторов головы, иннервируемых передним нервом боковой линии. Дорсальное ядро у этих животных отсутствует (рис. 35, б).
Возможным объяснением подобных различий в организации первого центрального отдела системы органов боковой линии является специализация ее в связи с электрорецепцией. Действительно, среди водных позвоночных обнаружены виды, чувствительные к электрическим воздействиям и воспринимающие низкочастотные и низкоамплитудные сигналы, которые распространены в водной среде. Электрочувствительными являются миноги, некоторые представители рыб и хвостатых амфибий. Отдельные виды рыб способны не только воспринимать, но и генерировать электрические разряды (так называемые электрогенные рыбы). Подобного рода особенности, а также происхождение элементов электросенсорных систем обусловили различия в характере организации центральных звеньев органов боковой линии.
Полагают, что дорсальное ядро октаво-латеральной системы является структурой, специфически связанной с электрорецепцией. Об этом же свидетельствует его присутствие в мозге электрочувствительных амфибий. Вместе с тем электрочувствительные костистые рыбы (клюворылые, гимнотовидные, сомообразные и араванообразные) лишены дорсального ядра. У них информация от электрорецепторов по системе нервов боковой линии достигает особой, электросенсорной, области, которая занимает латеральную часть акустико-латеральной зоны продолговатого мозга (рис. 28, д, е). В этой области обнаруживается ряд структурных изменений по сравнению с медиальной частью, куда проецируются механорецепторы органов боковой линии.
Так, под глубоким слоем мозжечкового гребня формируется плексиморфный слой, в котором располагаются дендриты клеток, подобных клеткам Пуркинье мозжечка. На них, а также на телах округлых клеток этого слоя заканчиваются аксоны первичных сенсорных нейронов. Крупные клетки являются проекционными, направляющими аксоны к вышележащим отделам, округлые клетки - комиссуральными.
У рыб, способных генерировать электрические сигналы, обнаруживается дальнейшая специализация области центрального представительства органов боковой линии. В медуллярном отделе значительно утолщается молекулярный слой, зона крупных клеток дифференцируется на пять самостоятельных слоев, число клеточных типов возрастает до десяти. При этом разные типы клеток получают информацию от разных видов рецепторов, а эффекторные элементы медуллярного уровня проецируются в разные отделы торуса среднего мозга (рис. 37). В ряде случаев у электрогенных рыб гипертрофированными оказываются система латеральной петли и торус среднего мозга (рис. 38).
Считается, что электрорецепция в эволюции появляется сравнительно рано - у животных, давших начало миногам и хрящевым рыбам. Однако, хотя она сохраняется во многих линиях позвоночных, включая предков наземных, в более молодых группах - у всех ныне живущих голостей и большинства
телеостей - она, по-видимому, была утрачена, что сопровождалось исчезновением дорсального ядра октаво-латеральной области. В настоящее время лишь у 1% костистых рыб имеются органы электрорецепции. При этом и
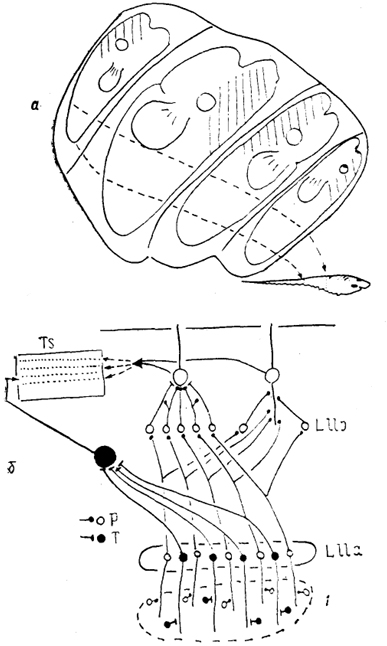
Рис, 37. Схема организации проекций электрорецепторов в мозге Gymnoti-formis (Maler e. a., 1981; Heiligenberg, Dye, 1982).
а - распределение входов в заднюю долю органов боковой линии от рецепторов разных отделов тела, б - представительство афферентов от двух типов бугорковых рецепторов (Р и Т) в стволе мозга. 1 - рецептивная поверхность.
рецепторная часть и область центральных проекций электросенсорной системы значительно отличаются от таковых у видов "основной" линии, что может свидетельствовать о ее независимом и позднем появлении в эволюции.
Таким образом, медуллярное представительство органов боковой линии у разных групп рыб (рыбы, не обладающие электрорецепцией, костистые рыбы с электросенсорной системой, электросенсорные рыбы, не относящиеся к костистым, и электрогенные
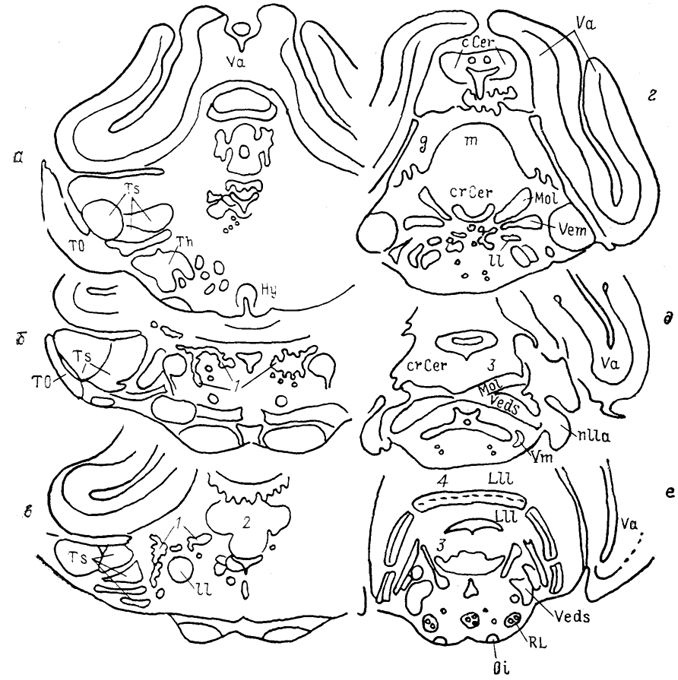
Рис. 38. Ядра ствола мозга представителя мормирид Gnathonemus petersi (Meek e. а., 1986),
a - e - срезы в рострокаудальном направлении. 1 - латеральное ядро заслонки мозжечка; 2 - гранулярное возвышение, 3, 4 - области проекций механо- (3) и электрорецепторов (4) в зоне представительства органов боковой линии.
рыбы) организовано по-разному, а электрорецепция и связанные с нею мозговые образования костистых рыб не гомологичны таковым у других групп животных.
У всех амфибий система органов боковой линии присутствует, по крайней мере в личиночной стадии. В периферическом отделе она включает механо-, а у некоторых и электрорецепторы. Центральная организация медуллярного представительства подобна той, которой обладают рыбы, не относящиеся к костистым. Ядра октаво-латеральной системы амфибий занимают перивентрикулярное положение.
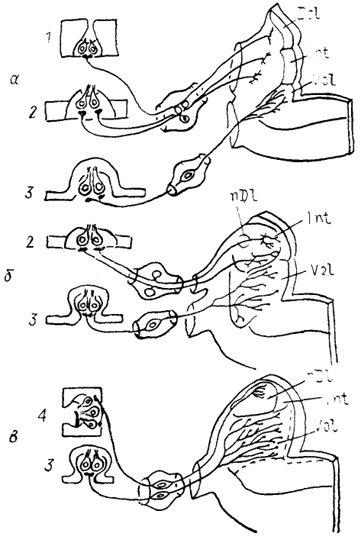
Рис. 39. Октаво-латеральная область продолговатого мозга амфибий (Fritzsch e. а., 1985).
а - электрочувствительные хвостатые амфибии, б - хвостатые и личинки бесхвостых, в - взрослые бесхвостые амфибии. 1-4 - рецепторы октаво-латеральной системы: электрорецепторы (1), механорецепторы органов боковой линии (2), вестибулярные (3), акустические (4).
У личинок всех амфибий и взрослых особей водных хвостатых амфибий передний нерв боковой линии имеет несколько корешков. Дорсальный корешок проводит информацию в область продолговатого мозга, названную дорсальным островком Кингсбери, или дорсальным ядром октаво-латеральной области, nucl. dorsalis areae octavolateralis. На основании томографического сходства и наличия входов от электрорецепторов его гомологизируют с дорсальным ядром примитивных электрорецептивных рыб. В зону, лежащую вентрально от него, входит пучок, проводящий информацию от механорецепторов органов боковой линии. Этот отдел-гомолог медиального ядра октаво-латеральной области рыб - носит название промежуточного ядра, nucl. intermedius areae octavolateralis (изначально описано как каудальное ядро), и у бесхвостых амфибий перед метаморфозом редуцируется до маленькой клеточной группы. По имеющимся данным оно получает небольшое число входов от вестибулорецепторов (рис. 39). У личинок бесхвостых амфибий имеется дорсальный островок Кингсбери, однако у взрослых особей, даже обитающих в воде, его нет, что связывают с отсутствием органов электрорецепции. Органы боковой линии, сохраняющиеся у некоторых бесхвостых амфибий, проецируются в промежуточное (медиальное) ядро. Восходящие связи медуллярного центра направляются в средний мозг. У некоторых безногих амфибий сохраняются и механо- и электросенсорные органы боковой линии, однако их центральные проекции недостаточно изучены.
Таким образом, при специализации органов боковой линии,
каждая из рецепторных систем раздельно представлена на медуллярном уровне, причем области представительства топологически четко организованы и часто имеют ламинарный характер строения.
Акустическая зона (октавная - у низших) продолговатого мозга включает в себя два комплекса ядер: ядра, расположенные дорсолатерально и связанные с обработкой информации об акустических сигналах и лежащие вентромедиально вестибулярные ядра. Обе группы составляют область представительства VIII черепномозгового нерва,* n. vestibulocochlearis, и занимают вентральное положение относительно центров боковой линии.
В мозге круглоротых система VIII нерва проецируется в вентральную часть октаво-латеральной области (рис. 29). В ее пределах некоторые авторы выделяют три ядра (дорсальное, медиальное, тангенциальное), другие - два (медиальное и заднее), но большинство склоняется к мысли о гомологичноети всей этой области вентральному ядру октаво-латеральной области рыб. Эфференты этого отдела направляются в спинной мозг и в составе медиального продольного пучка - к глазодвигательному комплексу. Имеются сведения о наличии проекций в торус, ядро задней комиссуры и диенцефальную область. Часть входов адресована ретикулярной формации.
В мозге рыб волокна VIII иерва проецируются в вентральную зону октаво-латеральной области продолговатого мозга. Конкретное описание ее разными авторами проводится по-разному и вопросы гомологизации ядер октавного комплекса далеко не разрешены.
Полагают, что вентральное ядро, получающее в мозге круглоротых входы от вестибулорецепторов и проецирующееся на спинной мозг, сохраняется и в мозге рыб (рис. 29, 35, 36). При этом происходит увеличение количества и отчасти размеров составляющих его элементов. У многих групп рыб (хрящевых, саркоптеригий и актиноптеригий за исключением телеостей) описывают четыре ядра системы VIII нерва - переднее, крупноклеточное,** нисходящее и заднее вестибулярные. У костистых рыб появляются дополнительные ядра этого комплекса: ядро Дейтерса и тангенциальное.
Эфференты октавной системы адресуются многим отделам мозга, в ,их числе вышележащие образования - торус, мозжечок, а также моторные ядра ствола. Связи с моторными ядрами образуют особый пучок, tr. octavomotorius, волокна которого проходят в составе иней- и контрлатерального медиальных продольных пучков и формируют восходящую и нисходящую
ветви. Восходящая ветвь направляется к ядрам III и IV нервов, нисходящая - к ядру VI нерва и мотонейронам спинного мозга. Волокна, идущие к спинному мозгу, у костистых рыб могут проходить параллельно медиальному продольному пучку. Среди них выделяют перекрещенный латеральный октаво-спинальный тракт, который, как полагают, берет начало от тангенциального ядра, и прямой медиальный октаво-спинальный тракт, идущий от крупноклеточного вестибулярного ядра.
Значительное число эфферентов после перекреста восходит рострально и направляется в торус среднего мозга. Эта система носит название латеральной петли, lemniscus lateralis, или .латерального продольного пучка, fasc. longitudinalis lateralis, и хорошо выражена у всех лучеперых рыб. Ее волокна, помимо торуса, адресуются латеральному ядру заслонки мозжечка, а у костистых рыб - tectum opticum.
Эфференты октавной части хрящевых рыб изучены недостаточно подробно. Имеются сведения о наличии проекций к спинному мозгу и мозжечку (рис. 36, А). У голостей и костистых рыб показано широкое и топографически организованное представительство разных участков вестибулярных ядер в различных отделах мозжечка.
Комиссуральные связи октавной области составлены 'внутренними дугообразными волокнами, которые проходят в области тегментума вентрально от медиального продольного пучка.
Для всех групп рыб обнаружена эфферентная иннервация октавной части. Источники связей локализованы в пределах ретикулярной формации и составляют "октаво-латеральное эфферентное ядро". У костистых рыб описана также дополнительная группа клеток, расположенная в промежуточном мозге.
В мозге амфибий основные входы волокон VIII нерва адресованы вентральному ядру акустико-латеральной области, а их коллатерали (у хвостатых и личинок бесхвостых) - латеральной части промежуточного ядра.* Кроме того, имеются входы и в латеральную зону ретикулярной формации. В составе вентрального вестибулярного ядра у разных видов описано от двух до пяти отделов (рис. 30). Его эфференты образуют октаво-моторный тракт, а само ядро (или его часть) рассматривается как гомолог ядра Дейтерса. Эфференты вестибулярных ядер амфибий формируют и нисходящие проекции к рецепторам вестибулярного аппарата.
В мозге рептилий вестибулорецепторы проецируются в крупноклеточное ядро, nucl. vestibularis magnocellularis, - возможный гомолог ядра Дейтерса - и лежащее латерально тангенциальное ядро, nucl. tangentialis. У многих рептилий, однако,
имеется более сложное развитие комплекса и в его составе выделяют до шести ядер (рис. 31). Крупноклеточное ядро (вентролатеральное вестибулярное по другой классификации) я его основной путь - латеральный вестибуло-спинальный тракт - достигают значительного развития у четвероногих рептилий, у безногих же особенно велико тангенциальное ядро.
Нейроны вестибулярного комплекса рептилий формируют два нисходящих тракта - латеральный и медиальный вестибуло-спинальные, trr. vestibulospinalis lateralis et medialis. Первый берет начало от крупноклеточной части вентролатерального и тангенциального ядер и идет ипсилатерально. Второй начинается от контрлатеральных вентромедиального, нисходящего, вентролатерального и от ипсилатерального дорсолатерального ядер. Восходящие проекции адресованы структурам среднего мозга (контрлатеральным ядрам III и IV нервов) и проходят в составе медиального продольного пучка. В ряде случаев доказано, что как восходящая, так и нисходящая ветви могут образовываться одним и тем же нейроном в результате бифуркации аксона. Такие клетки описаны в медиальном и дорсолатеральном вестибулярных ядрах.
В мозге птиц ядра вестибулярного комплекса сохраняют свойственные им состав и связи. Помимо них выделяют и дополнительные ядра - дорсомедиальное и верхнее вестибулярные (рис. 32). Эфференты вестибулярного комплекса у рептилий и птиц оказываются сходными и отличаются по степени выраженности. Основные проекции адресованы мозжечку, моторным ядрам черепномозговых нервов, ретикулярной формации, глазодвигательному комплексу. Имеются и билатеральные входы к рецепторным отделам. Среди вестибуло-спинальных лроекций выделяют латеральный и медиальный тракты. Первый проходит в латеральных канатиках, а второй - в латеральном отделе вентральных канатиков. Кроме того, по системе дугообразных волокон проходят связи с контрлатеральным вестибулярным комплексом. Помимо входов от нейронов ганглия Скарпа в вестибулярный комплекс рептилий и птиц поступают волокна спинно-вестибулярного тракта.
В вестибулярном комплексе млекопитающих значительно редуцируется тангенциальное ядро, а в составе латерального ядра обнаруживаются как крупные, так и мелкие нейроны. Каудальным продолжением мелкоклеточного отдела ядра Дейтерса является нижнее, или нисходящее, ядро. Медиальное ядро хорошо развито и его гомологи обнаруживаются в мозге рептилий и птиц. Кроме названных в составе вестибулярного комплекса представлено верхнее, или ядро Бехтерева, а также группы клеток X, Y, Z. Входы от нейронов ганглия Скарпа организованы таким образом, что информация от рецепторов полукружных каналов адресуется верхнему и медиальному ядрам, нижнее и латеральное получают входы от рецепторов саккулуса
и утрикулуса. Активация рецепторов саккулуса, кроме того, приводит к изменению активности нейронов группы Y, утрикулуса - медиального вестибулярного ядра. Помимо первичных сенсорных входов ядра комплекса получают проекции от разных отделов ретикулярной формации, мозжечка, спинного мозга. Весьма сложными оказываются связи между ядрами комплекса. Комиссуральные связи опосредуются в основном нейронами группы Y.
Эфференты вестибулярных ядер направляются к моторным структурам, мозжечку, ретикулярной формации в составе медиального продольного пучка, вестибуло-спинального и вестибуло-мозжечкового трактов. Часть волокон адресуется ядрам глазодвигательного комплекса (III, IV и VI черепномозговых нервов). Нейроны латерального вестибулярного ядра осуществляют иннервацию рецепториого отдела.
У млекопитающих система проекций вестибулярных ядер значительно усложняется в связи с развитием тонких зрительно-моторных координации. При этом остается открытым вопрос о наличии и функциональной роли проекций вестибулярной системы, восходящих к таламо-теленцефальному уровню.
В целом у позвоночных обнаруживается удивительное сходство в характере связей вестибулярной системы: многочисленные проекции к исполнительным отделам мозга определяют решающую роль данной сенсорной системы во взаимодействии со зрительной, проприоцептивной и соматической системами и обеспечивают осуществление позных рефлексов и ориентацию в трехмерном пространстве.
Наиболее сложной, по-видимому, является проблема эволюционного становления акустической части октаво-латеральной системы, здесь остаются нерешенными вопросы как возникновения, так и гомологизации слуховых центров. Так, неясно, имеют ли бесчелюстные в дополнение к вестибулярным слуховые рецепторы и соответствующие отделы в продолговатом мозге. Считается, что рыбы уже обладают слуховыми рецепторами внутреннего уха, однако отдельного центрального их представительства не обнаружено. Вместе с тем для одной группы рыб показано, что наряду с четко организованными входами вестибулорецепторов в определенные области октавного комплекса проецируются и рецепторы саккулуса, возможно связанные с восприятием акустических стимулов. Это позволяет предположить, что, по крайней мере, некоторые группы рыб, подобно тетраподам, имеют раздельные акустические и вестибулярные области продолговатого мозга и соответственно ветви VIII нерва.* Однако вряд ли это является правилом для организации
слуховой системы всех рыб, поскольку рецепторы внутреннего уха у них оказываются гетеромодальными и могут быть активированы как акустическими, так и гравитационными воздействиями. Более того, у разных рыб функцию восприятия акустических раздражителей выполняют разные периферические отделы октавной системы.
Особенно сложен вопрос об организации слуховых центров и их происхождении у амфибий, поскольку именно на этом уровне появляются четка дифференцированные мозговые структуры данной системы. Согласно хорошо известной гипотезе О. Ларселла у амфибий с исчезновением механорецептивных органов боковой линии промежуточное (медиальное) ядро становится центральным представительством слуховой, системы, преобразуясь в кохлеарное, или улитковое, ядро. Однако многочисленные исследования последних лет убедительно опровергли это мнение. Так, у бесхвостых амфибий акустические входы проецируются как в промежуточное, так и в дорсолатеральное ядро октаво-латеральной области продолговатого мозга (рис. 39). Последнее появляется непосредственно перед метаморфозом и становится основным медуллярным отделом слуховой системы. На месте промежуточного ядра у бесхвостых амфибий остается небольшая группа клеток, которая получает коллатерали слухового нерва. У хвостатых амфибий дорсолатеральное ядро отсутствует и акустический вход адресуется промежуточному ядру. Вместе с тем существует множество вариаций этой схемы. Так, у некоторых хвостатых амфибий, развивающихся без метаморфоза, промежуточное ядро не обнаружено. У тех безногих амфибий, у которых доминирует акустический вход, специфические слуховые образования отсутствуют вовсе. Все это позволило ряду авторов высказать предположение о многообразии путей становления слуховой системы в группе амфибий и, таким образом, допустить возможность параллельного развития слуховых образований в разных группах земноводных.
Представляется неясным происхождение акустического центра бесхвостых амфибий - дорсолатерального ядра (оно ранее описано под разными названиями, наиболее известное из которых "дорсальное крупноклеточное ядро". Гипотеза о происхождении его из медиального (механорецептивного в системе органов боковой линии) ядра рыб опровергается, по-видимому, наличием и промежуточного, и дорсолатерального ядер одновременно у тех бесхвостых амфибий, у которых во взрослом состоянии сохраняются органы боковой линии.
Другая линия доказательств независимости возникновения акустического ядра от редукции органов боковой линии проводится исходя из различий в характере эфферентных проекций этих систем. Проекции медуллярных центров адресованы разным отделам торуса, прямые связи слухового ядра с тектумом отсутствуют, наконец, в пределах продолговатого мозга бесхвостых амфибий формируется новое скопление клеток, получающее входы только от слухового ядра,-верхняя олива, oliva superior (рис. 30, 5). Не получает подтверждений и представление о развитии акустического центра из дорсального ядра октаво-латеральной области. Действительно, потеря электрорецептивных
органов и появление дорсолатерального ядра, занимающего положение, идентичное положению дорсального (электросенсорного) ядра, совпадают во времени. Исходя из этого было высказано мнение о трансформации данной структуры. Однако изучение генеза нейронов слухового ядра опровергает эти предположения: его источники, по-видимому, локализуются и в вентральной, и в дорсальной частях акустико-латеральной области. Последняя представлена пролиферативной зоной, которая после утраты дорсального ядра дает начало клеткам акустического ядра. Вентральная же, вероятно, гомологична области, занимаемой в мозге рыб вентральным ядром. Об этом свидетельствует сходный характер связей.
Эфференты дорсолатерального ядра направляются к контрлатеральному одноименному образованию, в торус среднего мозга и билатерально в верхнюю оливу, которая располагается, в пределах медуллярной ретикулярной формации и в оформленном виде появляется у личинок бесхвостых амфибий перед, метаморфозом. Слуховое ядро составляет основной объем ее афферентов. Эфференты верхней оливы, а также некоторое число волокон слухового ядра составляют латеральный бульбо-тектальный тракт, или латеральную петлю, и адресуются торусу. Рассеянные среди волокон клеточные тела рассматриваются как предшественники ядер латеральной петли высших, nucll. lemnisci lateralis.
В мозге разных рептилий акустический комплекс обнаруживает значительное число вариаций строения. Так, у черепах, многих ящериц, амфисбен в составе этой области выделяют два ядра, нейроны которых получают входы слуховой ветви-VIII нерва. Ядра имеют разные названия, чаще всего их обозначают как переднее и заднее кохлеарные (улитковые), nucll. cochlearis anterior et posterior. У змей описана лишь одна клеточная группа. Значительно сложнее устроен кохлеарный комплекс у некоторых ящериц (варан, хамелеон, игуана, агама), и особенно у крокодилов. У последних состав и характер связей сходны с имеющимися у птиц. У названных рептилий-в пределах переднего гипертрофированного ядра выделяют ростральное, угловое и дорсальное крупноклеточное кохлеарные ядра, nucl. cochlearis rostralis, angularis et dorsalis magnocellularis соответственно (рис. 31). Полагают, что дорсальное крупноклеточное ядро рептилий - гомолог акустического - ядра амфибий и вентрального кохлеарного ядра млекопитающих, угловое же, не имеющее предшественников в мозге бесхвостых амфибий, превращается у млекопитающих в дорсальное кохлеарное ядро. Эфференты дорсального крупноклеточного ядра направляются к ламинарному ядру, которое рассматривается как гомолог медиальной верхней оливы и, возможно, ядер трапециевидного тела. Восходящие проекции всех слуховых
ядер направляются к верхней оливе и в слуховые центры среднего мозга.*
В мозге птиц слуховые центры медуллярного отдела занимают большой объем и имеют сложное строение (рис. 32). Ядра кохлеарного комплекса состоят из дорсального и вентрального ядер. Они рассматриваются как гомологи соответственно углового и крупноклеточного ядер рептилий. От вентрального ядра проекции адресуются ламинарному ядру, nucl. cochlearis laminaris, которое на основании сходства связей считают гомологичным медиальлой верхней оливе млекопитающих-Нейроны углового и ламинарного ядер дают начало крупному пучку волокон (возможно, трапециевидному телу), направленному контрлатерально и передающему информацию верхней оливе и ядрам латеральной петли противоположной части мозга. Часть волокон восходит к среднему мозгу. Наконец, некоторая часть проекций кохлеарных ядер птиц адресована ретикулярной формации и моторным ядрам черепномозговых нервов.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|