- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 8 страница
У птиц каудальные 2/3 ядра XII нерва, nucl. n. hypoglossi Iracheosyringealis, содержат мотонейроны, осуществляющие иннервацию голосового аппарата. Входы в эту область образованы структурами среднего мозга и перешейка, связанными с обеспечением вокализации.
Ядро XII нерва у млекопитающих подразделяют на дорсальную и вентральную части, обеспечивающие движения языка в результате иннервации разных мышц. Афференты к нему поступают от главного ядра V нерва, вентрального парабрахиального и ядра Колликера-Фузе. У хищных, в отличие от крыс и опоссума, выражены проекции мезенцефалического ядра
V нерва и прямые сенсорные входы от клеток яремного ганглия, проводящих информацию от проприорецепторов. Часть афферентов поступает на дендриты нейронов ядра, проникающие в пределы ретикулярной формации (медиальной зоны), нисходящего ядра V нерва, ядра солитарного тракта и, возможно, мозжечка, поскольку некоторые дендриты распределяются среди волокон медиального продольного пучка.
Часто в составе ядра XII нерва у высших позвоночных описывают предлежащее ядро, nucl. praepositus hypoglossi. Однако по характеру связей оно существенно отличается от основной части. К настоящему времени и для млекопитающих, и для рептилий показаны афференгы, поступающие к нему из спинного мозга и вестибулярного комплекса. Эфференты предлежащего ядра направляются в мозжечок, спинной мозг, комплекс глазодвигательных ядер, чем обусловлено участие этого ядра в организации движений глаз. Ряд авторов полагает, что эта структура является вентральным продолжением вестибулярного комплекса, но она лишена первичных сенсорных входов.
Таким образом, на уровне продолговатого мозга позвоночных находится основной объем ядер черепномозговых нервов. В этом отношении медуллярный отдел головного мозга имеет множество общих черт строения и сходство в организации связей у представителей разных групп. Однако, наряду с этим, при сравнительноанатомическом изучении выявляется постепенное усложнение структурной организации, обусловленное целым рядом факторов и, прежде всего, прогрессивной эволюцией нервной системы в целом и адаптивной эволюцией, приводящей к специализации определенных систем.
Результатами эволюционных преобразований в нервной системе, в частности, являются изменение взаимосвязей продолговатого мозга с другими отделами нервной системы (что выражается в усложнении системы проводящих путей) и снижение удельного веса ядер черепномозговых нервов в общем объеме структур данного отдела. Последнее есть следствие общего последовательного усложнения взаимоотношений в нервной системе, в результате чего многие образования разных уровней оказываются включенными в состав какой-либо определенной системы мозга. В этом случае речь идет об участках мозга, которые невозможно отнести к сенсорным или моторным образованиям. Их функциональная роль может быть понята только из анализа деятельности той системы, в состав которой они включены. Естественно, что такие структуры в основном принадлежат вышележащим отделам мозга, однако часть из них обнаруживается и в пределах ромбенцефалона. К ним относятся нижняя олива, собственные ядра моста, дугообразное ядро и некоторые другие, представляющие собой звенья моторных систем мозга. Развитие и совершенствование последних приводит
к преобразованию названных медуллярных ядер в мозге высших позвоночных. У низших гомологи или аналоги подобных ядер обнаруживаются зачастую в пределах ретикулярной формации (см. гл. 6).
* Так, например, в доле блуждающего нерва карпообразных верхние слои являются сенсорными, а нижние содержат моторные элементы.
* Спинно-окципитальные нервы присутствуют в мозге многих актиноптеригий и занимают положение между бранхиальными и спинальными нервами. Они состоят из двух групп волокон: окципитальных, выходящих из полости черепа, и окципито-спинальных, место выхода которых располагается сразу каудально от черепа. Число этих нервов варьирует. У некоторых телеостей они отсутствуют.
§ 4. ЯДРА ОБЛАСТИ ПОКРЫШКИ ПРОДОЛГОВАТОГО МОЗГА
Помимо ядер, относящихся к системе черепномозговых нервов, на уровне продолговатого мозга сосредоточен и ряд других образований, в том числе нижняя олива, oliva inferior, - типичное производное ретикулярной формации, представляющее собой совокупность интернейронов. Ее выраженность связана со степенью развития мозжечка и моторных систем мозга.
Некоторые авторы описывают нижнюю оливу уже в мозге миног, где эта совокупность клеток, лежащих в пределах ретикулярной формации, отличается от соседних характером связей. Это образование получает входы от дорсальных отделов спинного мозга, тектума и той зоны ретикулярной формации, которую рассматривают как прекурсор красного ядра. Однако вычленение этой группы клеток из окружающих ретикулярных производных весьма затруднительно, и этим объясняется значительное число противоречий в описаниях нижней оливы у низших позвоночных.
В мозге акул нижняя олива (или ее прекурсор) представлена группой мелких клеток, лежащих парамедианно в каудальной части продолговатого мозга (рис. 34). Многие авторы, не убежденные в ее гомологичности нижней оливе высших позвоночных, предпочитают использовать другие названия: "ядро А", "парасептальное ядро". Тем не менее, показано, что от данной группы клеток берет начало путь, адресованный мозжечку (возможный гомолог оливо-мозжечкового тракта). Кроме того, на этот нейронный комплекс проецируются структуры конечного мозга.
Небольшое по размерам ядро нижней оливы обнаружено у представителей всех групп актиноптеригий. Оно лежит вентрально в каудомедиальной части ромбовидного мозга (рис. 35) и состоит из мелких нейронов, ориентированных параллельно поверхности. Афференты ее образованы спинным мозгом, а у костистых рыб - и вышележащими отделами. Эфференты нижней оливы полностью контрлатеральны и топологически строго распределяются в мозжечке, формируя характерные лиановидные волокна.
Строение нижней оливы значительно варьирует, и для многих амфибий и рептилий ее описания отсутствуют. У некоторых рептилий нижняя олива представлена небольшой диффузной группой клеток, которую трудно отделить от ретикулярной формации. Тем не менее у хамелеонов и крокодилов это
уже крупное скопление клеток, сходное с таковым у птиц. Кроме того, для многих рептилий характерно наличие значительного числа лиановидных волокон в мозжечке, хотя их оливарное происхождение экспериментально не доказано.
У птиц нижнеоливарный комплекс подразделяется на дорсальную и вентральную оливы (рис. 32). Первая в свою очередь делится на медиальную и латеральную части, или ламеллы. Полагают, что медиальная ламелла птиц соответствует нижней оливе рыб и наиболее выражена у водных позвоночных. Латеральную ламеллу иногда рассматривают как гомолог нижней оливы крокодилов. Дорсальная олива у большинства птиц превосходит вентральную, - гомолог главного ядра нижней оливы млекопитающих. Эфференты нижней оливы у птиц, закономерным образом проецируются на разные участки поверхности мозжечка, формируя типичные лиановидные волокна.
Нижняя олива млекопитающих - ядерный комплекс, лежащий в основании продолговатого мозга латерально от волокон пирамидного тракта. В него входят главное ядро нижней оливы, наибольшее по размерам и весьма характерной формы, а также медиальная и дорсальная добавочные оливы. Полагают, что первая гомологична медиальной, а вторая латеральной ламеллам нижней оливы птиц.
Наибольшей сложности этот комплекс достигает у антропоидов, у которых сильно развитое главное ядро обусловливает появление на вентролатеральной поверхности продолговатого мозга одноименных выпуклых образований - олив, olivae. Они отсутствуют у однопроходных, сумчатых, трубкозубых, грызунов. Среди хищных оливы продолговатого мозга хорошо выражены у панд, медведей, менее - у кошек. Весьма развиты оливы у китов.
Для представителей других групп млекопитающих характерно преобладание добавочных олив, хотя их развитие варьирует в широких пределах. Так, у крыланов медиальная добавочная олива подразделяется на несколько клеточных групп, а у ондатры - это фактически единственная хорошо развитая часть нижнеоливарного комплекса.
Афференты добавочных олив сформированы спинномозговыми волокнами, ядрами дорсальных канатиков, медиальным и промежуточным ядрами мозжечка. Главное ядро получает проекции от красного ядра, многочисленных мелких ядер среднего мозга и контрлатерального зубчатого ядра мозжечка; все ядра комплекса - от коры большого мозга.
Эфференты всех ядер нижней оливы образуют контр латеральные входы в мозжечок, поступая через нижние ножки и распределяясь в виде парасагиттально ориентированных полос в коре мозжечка. Здесь они получают название "лиановидные волокна" и контактируют с дендритами клеток Пуркинье.
Таким образом, на уровне продолговатого мозга располагаются структуры, выполняющие релейные функции - обеспечение взаимосвязи высших отделов мозга и мозжечка. Помимо нижней оливы у птиц и млекопитающих обнаруживается особое скопление клеток с аналогичными функциями - ядра моста, nucll. pontis, которые не описаны даже в мозге рептилий.
У птиц они состоят из двух клеточных групп, расположенных в вентральной части ромбовидного мозга - от уровня моторного ядра V нерва рострально до комплекса ядер VIII нерва каудально. Аксонами одной из комиссур мозга они отделены от окружающей ретикулярной формации. Кроме того, составляющие их клетки отличаются от ретикулярных как формой, так и размерами. Латеральная клеточная группа расположена рострально. Медиальная имеет более четкие границы и смещена каудально. Нейронный состав их идентичен. Обе группы проецируются контрлатерально в кору мозжечка, в латеральные отделы долек VI-VIII.
У млекопитающих развитие подобных нейронов и их сосредоточение в области покрышки приводит к формированию самостоятельного отдела ромбовидного мозга - моста, Pons. Его выраженность зависит от степени развития мозжечка и коры большого мозга: у утконоса и некоторых рукокрылых нейроны ядер моста не образуют крупных скоплений и распределены среди других компонентов покрышки, у сумчатых их дорсальную границу составляют ножки мозга. Крупное скопление мостовых ядер обнаруживается у человека, человекообразных обезьян и дельфинов. В этих случаях волокна ножек мозга, разделяясь на небольшие пучки, распределяются среди мостовых ядер. Сильным развитием характеризуются они и у слонов, но в этом случае пирамидный тракт, не пронизывая их, проходит дорсально.
Афферентами ядер моста служат входы различных областей новой коры. Эфференты дают начало средним ножкам мозжечка, волокна которых адресованы коре мозжечка. В средних ножках проходит и часть церебелло-понтийных волокон.
Естественно, что наличие столь обширного скопления клеток значительно меняет топографические отношения и внешний вид ромбовидного мозга млекопитающих. В результате часть ядер, расположенных у всех позвоночных в пределах продолговатого мозга, у млекопитающих мигрирует рострально и оказывается включенной в понтийный отдел (главное ядро V нерва, ядро отводящего нерва и др.).*
Часть нейронов, отделяясь от ядер моста, у млекопитающих мигрирует каудально и образует в вентральных отделах продолговатого
мозга дугообразные ядра, nucll. arcuati, которые сохраняют свойственные понтийным ядрам связи.
В продолговатом мозге низших позвоночных в пределах тегментума можно выделить так называемые премоторные образования, обеспечивающие иннервацию сегментарных моторных ядер. В основном они представляют собой разные отделы ретикулярной формации, однако в ряде случаев образуют дискретные структуры, легко вычленяемые из ретикулярного окружения. К образованиям подобного рода относятся гигантские клетки Мюллера и Маутнера, типичные для мозга низших позвоночных (рис. 26).
В мозге круглоротых мюллеровские клетки образуют три дискретные группы - в области продолговатого мозга, перешейка и среднего мозга. Их аксоны вместе с аксонами маутнеровских клеток в составе медиального продольного пучка направляются к моторным отделам спинного мозга. Сведения относительно существования маутнеровских клеток у хрящевых рыб противоречивы; некоторые авторы описывают их в мозге акул. Аксоны этих клеток, как и у других позвоночных, адресованы спинному мозгу, куда они проходят в составе медиального продольного пучка.
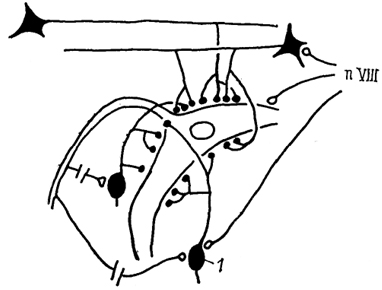
Рис. 43. Схема связей маутнеровской клетки костистых рыб (Faber e. а., 1989).
1 - коллатеральный нейрон. Светлые кружки - возбуждающие синапсы, темные - тормозные.
У актиноптеригий маутнеровские клетки лежат в латеральной части базальной пластинки ромбовидного мозга на уровне вхождения VIII черепномозгового нерва (рис. 43). Каждая из клеток обладает двумя главными дендритами, один из которых простирается вентрально, а другой - латерально. Кроме того, имеются короткие дендриты, берущие начало от тела клетки. Латеральные дендриты распределяются в вентральной части октаво-латеральной области, где получают возбуждающий вход от волокон VIII нерва. Помимо возбуждающих входов от нервов боковой линии и VIII нерва маутнеровокие клетки получают тормозные входы от клеток, лежащих в вентральной части контрлатеральной октаво-латеральной зоны. Их аксоны образуют густую сеть в области начального сегмента аксона маутнеровской клетки. Кроме названных имеются проекции от главного ядра V нерва, мозжечка, тектума.
Последний опосредует поступление информации от зрительной системы, возможно, что эта афферентация адресуется вентральному дендриту, ветви которого распространяются в поверхностной зоне покрышки, т. е. в области, где проходят прямые и перекрещенные волокна текто-бульбарного тракта.
Сильно миелинизированные аксоны маутнеровских клеток направляются медиально, затем поворачивают каудально и входят в контр латеральный медиальный продольный пучок, который проходит по вентральным канатикам спинного мозга до его каудальных сегментов к интер- и мотонейронам. Активация подобной системы связей приводит к специфической моторной реакции - сильному удару хвоста - возникающей с коротким латентным периодом в результате сокращения мышц, контрлатеральных стороне воздействия адекватных стимулов (зрительных, слуховых, а также активирующих органы боковой линии).
Маутнеровские клетки описаны и в мозге хвостатых амфибий, где они располагаются вблизи крупноклеточного вестибулярного ядра, а их латеральные дендриты ветвятся в области вхождения волокон октавного нерва. Отметим, что на основании расположения в мозге и тесных связей со структурами октаво-латеральной системы маутнеровские клетки иногда включают в состав последней.
Специализированные премоторные структуры обнаруживаются у животных, обладающих электрическими органами. У пластиножаберных (ромботелые и электрические скаты) электрические органы иннервируются электромотонейронами спинного или ромбовидного мозга. Во всех случаях афференты к ним сформированы аксонами нейронов, сосредоточенных в пределах нижнего ядра шва в вентральной части тегментума. У костистых рыб, обладающих электрическими органами мышечного происхождения, иннервация их осуществляется с медуллярного уровня особой группой клеток, образующих так называемое медуллярное пейсмекерное (командное) ядро. В его составе выделены крупные н мелкие нейроны. Последние, вероятно, представляют собой интернейроны, крупные же являются мотонейронами. Распределение клеток в пределах ядра у разных представителей варьирует от диффузного до топографически четко разделенного. Эфференты ядра адресованы электромотонейронам, лежащим в спинном мозге. Отметим, что у всех исследованных видов обнаружен топологический характер распределения проекций. Вопрос о гомологичности пейсмекерного ядра костистых рыб аналогичному образованию хрящевых остается невыясненным. Существует ряд отличий как в клеточном составе ядер, так и в характере внутриядерных взаимодействий. Возможно, с учетом предполагаемой независимости возникновения электрических органов у разных групп рыб,
речь идет об аналогичных функциях самостоятельных отделов мозга.
Таким образом, ромбовидный мозг позвоночных содержит структурные элементы разных сенсорных систем, а также моторные ядра черепномозговых нервов, определяющие организацию многих приспособительных реакций. Значительный объем ромбовидного мозга составлен структурами ретикулярной формации, которые также организуют проведение информации к моторным сегментарным отделам. В строении ромбовидного мозга весьма ярко отражаются те черты специализации, которые приобретают животные в свойственной им среде обитания. При этом экологически обусловленные модификации внешнего строения оказываются зачастую более широкими, нежели связанные с общим уровнем филогенетического развития. У низших позвоночных в области ромбовидного мозга и перешейка располагаются структуры, гомологи которых у млекопитающих лежат мезенцефально (их описание будет дано в гл. 5).
* Анатомия продолговатого мозга у млекопитающих изменяется и в результате появления пирамидного тракта, составляющего вентральную часть продолговатого мозга этих животных.
ГЛАВА 4
МОЗЖЕЧОК
Мозжечок, Cerebellum, всех позвоночных происходит в филогенезе из билатерального расширения, занимающего ростральные отделы ромбовидного мозга. Сначала связь между этими отделами осуществляется мембраной крыши IV желудочка. Позже эти парные образования смыкаются по средней линии своими ростральными участками, ромбовидная ямка расширяется и, наконец, клетки располагаются перпендикулярно оси мозга. Образовавшаяся пластинка формирует крышу ростральной части IV желудочка. Передняя ее часть связана тонкой пластинкой со средним мозгом, нижняя - образована мозговым парусом. Каудально примордиум мозжечка истончается в пластинку, служащую крышей IV желудочка.
У многих водных форм латеральные части мозжечка эвагинируют ростро-латерально, образуя так называемые ушковидные доли, или аурикулы, мозжечка, auriculae cerebelli (см. рис; 5). У челюстноротых медиальная часть мозжечка становится массивным телом мозжечка, corpus cerebelli. Его форма значительно варьирует, и различные отделы развиваются в результате разных процессов - инвагинации, эвагинации, утолщения стенок. Традиционно полагают, что аурикулы формируются под влиянием импульсации от образований октаво-латеральной системы, а тело - под воздействием соматосенсорных афферентов. Действительно, аурикулярные доли особенно хорошо развиты у животных, обладающих органами боковой линии: рыбы, хвостатые и бесхвостые амфибии (у последних в большинстве случаев в личиночной стадии). У амниот органы боковой линии отсутствуют, и аурикулы получают лишь вестибулярный вход, превращаясь в относительно небольшую часть мозжечка - клочково-узелковую долю, lobus flocculonodularis.
Тело мозжечка, увеличиваясь в размерах, покрывается бороздами, делящими его на доли и дольки. Его клетки сосредоточиваются у поверхности, образуя кору мозжечка. В глубине мозжечка уже у низших позвоночных выделяется группа клеток,
аксоны которых, выходя за пределы мозжечка, формируют систему его эфферентов. Совокупность этих клеток заметно возрастает в ряду позвоночных. У рыб и амфибий такие элементы либо диффузно распределены в толще мозжечка, либо объединены в небольшое ядро. У рептилий образуется два, а у птиц и млекопитающих три и более ядер. При этом они перемещаются к средней линии и приобретают значительные размеры.
Удивительно сходен и клеточный состав мозжечка позвоночных. Уже в мозге круглоротых можно выделить элементы, являющиеся прекурсорами основных типов нейронов, характерных для данного образования, - гранулярных нейронов и клеток Пуркинье. У высших позвоночных состав интернейронов мозжечка оказывается также сходным.
§ 1. МОЗЖЕЧОК НИЗШИХ ПОЗВОНОЧНЫХ
Наиболее просто устроен мозжечок круглоротых. Этот отдел мозга представлен у них пластинкой, содержащей узкий перивентрикулярный слой клеток, покрытый широкой наружной зоной нейропиля - молекулярным слоем. В клеточном слое содержатся тела мелких и крупных нейронов. Первые - предшественники гранулярных клеток мозжечка - обладают ориентированными к центру дендритами и латерально направленным аксоном. Дендриты крупных клеток простираются в молекулярный слой, аксоны направлены вентромедиально. Крупные клетки еще не обладают специфической особенностью клеток Пуркинье - богатым дендритным ветвлением, распространяющимся преимущественно в одной плоскости, и рассеяны среди мелких клеток. Среди афферентов преобладают входы от октаво-латеральной системы: сенсорные входы в составе нервов боковой линии и VIII черепномозгового нерва и проекции ядер октаво-латеральной области продолговатого мозга. В качестве других их источников описаны ядра тройничного нерва, тектум среднего мозга, гипоталамус.
Эфференты сформированы аксонами "клеток Пуркинье" и направляются в составе небольшого пучка к стволу, где распределяются среди структур продолговатого мозга, присоединяясь к медиальному продольному пучку. Ростральные проекции адресованы ретикулярной формации среднего мозга и ядру глазодвигательного нерва.
У взрослых миксин имеется совокупность волокон и клеток, напоминающая мозжечок других позвоночных. У Bdellostoma клеточный гребень, находящийся в той области, которую занимает мозжечок высших, внедряется в переднюю крышу ромбовидного мозга. У Ichthyomyzon и Entosphenus мозжечковый валик представлен пластинкой, лежащей между латеральными мозжечковыми дольками, которые в свою очередь являются продолжением
октаво-латеральной области продолговатого мозга. У миноговых латеральные дольки малы, а тело мозжечка представляет собой компактную клеточную массу, лежащую в перивентрикулярной области, где она формирует глубокую часть дорсолатеральной стенки IV желудочка и простирается рострально и медиально под фиброзной и молекулярной частями мозжечкового валика. Молекулярный слой - продолжение мозжечкового гребня, пересекая среднюю линию вместе с комиссурами, формирует тонкую ростральную зону, где проходят аксоны мелких нейронов латеральных долек и тела мозжечка. Присутствие тройничных и спинно-бульбарных волокон внутри тела и конвергенция входов от органов боковой линии и волокон VIII нерва позволили предположить, что молекулярный слой получает комплекс стимулов, отличающийся от адресованного мозжечковому гребню. Зона мозжечковых ядер представлена диффузной клеточной группой, которая, сохраняет топографическую близость с компонентами переднего октавного ядра.
У рыб размеры мозжечка, его положение, степень развития отдельных компонентов значительно варьируют не только в классах и отрядах, но и в пределах отрядов. В случае сильного развития мозжечка в нем выделяют центрально расположенное тело, два аурикулярных компонента и, у лучеперых, специфический отдел-заслонку мозжечка, valvula cerebelli. Хорошо развитые аурикулярные компоненты свойственны мозгу акул, у костистых рыб, напротив, особенно хорошо развита заслонка мозжечка при относительно слабом развитии ушковидных долей (см. рис. 27).
У хрящевых рыб мозжечок значительно более развит и дифференцирован, чем у бесчелюстных (см. рис. 5). На ранних стадиях их эмбриогенеза он, как и у миног, представлен пластинкой; на более поздних стадиях начинаются два независимых процесса: ростро-латерально направленное удлинение каудолатеральной части мозжечковой пластинки (приводящее к образованию аурикул) и дорсально направленная эвагинация ростро-медиальной части мозжечковой пластинки (дающая начало закладке тела мозжечка).
У взрослых пластиножаберных мозжечок состоит из тела и ушковидных долей. Последние представлены дорсомедиальным верхним листком и вентролатеральным нижним, продолжающимся в акустико-латеральную область. Тело мозжечка у многих представителей разделено бороздами на доли.
Степень развития мозжечка у хрящевых рыб весьма различна. Ранее полагали, что чем больше размеры животного, тем выраженнее деление тела мозжечка на доли. Так, у примитивных акул и химер поверхность мозжечка гладкая. В то же время у большинства акул и скатов его тело разделено поперечной бороздой, fissura prima. У более крупных видов появляются
и дополнительные борозды. Вместе с тем у животных одинакового размера можно обнаружить все вариации формы и степени развития мозжечка, что свидетельствует о возможности независимого происхождения борозд в разных группах. Таким образом, внешний вид мозжечка хрящевых рыб весьма разнообразен: химеры и катранообразные акулы, вероятно, обладают предковым типом мозжечка - гладким и разделенным на переднюю и заднюю доли. У других акул он выпуклый, сложно организованный и поделен на три доли. Сходные процессы независимо происходят у скатов, среди которых можно встретить представителей как с простым плоским мозжечком, так и сложным, изрезанным. У животных со вторично измененной билатеральной симметрией мозжечок асимметричный и имеет многочисленные борозды на поверхности.
Полость мозжечка, ventriculus cerebelli, невелика у скатов и значительна у акул. Основание тела мозжечка формирует ножку, brachium conjunctivum, которая соединяет ростральную часть со средним мозгом. Базальную часть ножки занимает так называемое вентральное возвышение мозжечка, внедренное в полость желудочка и составленное диффузно лежащими нейронами - ядро мозжечка, nucl. cerebelli, или латеральное ядро (см. рис. 34).
Тело мозжечка пластинюжаберных содержит четыре слоя: наружный молекулярный, str. moleculare externum; ганглионарный, str. ganglionare, содержащий клетки Пуркинье; волокнистый, str. fibrosum; внутренний гранулярный, str. granulare internum. Кроме того, гранулярные клетки образуют два продольных гребня - гранулярные возвышения, eminentiae granulares, которые, начинаясь сразу за местом перекреста блокового нерва, проходят вблизи средней линии по всей длине мозжечка (см. рис. 34).
Строение гранулярных клеток (или клеток-зерен) пластиножаберных подобно имеющемуся у всех позвоночных. Это плотно лежащие, мелкие клетки, обладающие тремя-шестью короткими дендритами. Последние один-два раза делятся и формируют характерного вида конечные ветвления. Аксоны, берущие начало от сомы или проксимального участка одного из дендритов, восходят в молекулярный слой и, Т-образно делясь, образуют систему параллельных волокон, идущих перпендикулярно оси мюзга. В гранулярных возвышениях пластиножаберных среди клеток-зерен обнаруживаются и более крупные элементы, подобные клеткам Гольджи. В латеральной части тела мозжечка гранулярных клеток нет, а имеется лишь фиброзная зона, содержащая афференты мозжечка и аксоны клеток Пуркинье.
Клетки Пуркинье распределяются почти по всему телу мозжечка за исключением парамедианной области (кроме Hydrotagus, у которых они распределены в молекулярном слое).
Клетки Пуркинье имеют форму, типичную для этих нейронов всех позвоночных: от удлиненной сомы отходит несколько крупных дендритов, которые, разветвляясь, формируют большое дендритное поле, уплощенное в парасагиттальном направлении. Ветви второго и более высоких порядков несут шипики.
Молекулярный слой представлен совокупностью параллельных волокон, дендритов клеток Пуркинье и волокон, подобных лиановидным высших позвоночных. В этом же слое присутствуют звездчатые интернейроны двух типов: поверхностно лежащие короткоаксонные и более глубоко залегающие клетки, аксоны которых оканчиваются на клетках Пуркинье, но, в отличие от корзинчатых клеток высших позвоночных, не имеют типичных для них терминалей.
В состав аурикул мозжечка акул входят три компонента: парные верхний и нижний листки и непарная срединная часть, pars medialis. или нижняя губа. Последняя составляет крышу IV желудочка (см. рис. 34, А). Все отделы различаются по структуре и характеру связей.
В аурикулах также выделяют молекулярный, ганглионарный и гранулярный слои. Фиброзный слой практически не выражен. Однако в каждом отделе аурикул есть особенности. Срединная часть имеет наиболее типичное строение и характеризуется хорошо выраженным ганглионарным слоем (кроме Hydrolagus), в котором дендритное поле клеток Пуркинье ориентировано перпендикулярно оси мозга (их направление меняется на сагиттальное лишь в области перехода в тело мозжечка). Гранулярные клетки помимо слоя, подстилающего клетки Пуркинье, образуют в верхнем листке аурикул парные латеральные и медиальные гранулярные возвышения - продолжение одноименных образований тела мозжечка. Их отростки формируют параллельные волокна мозжечкового гребня (хорошо развитого у акул). Кроме гранулярных в верхнем листке присутствуют клетки Гольджи. Нижний листок аурикул содержит незначительное число клеток Пуркинье, дендритное поле которых ориентировано перпендикулярно оси мозга.
В мозжечке скатов данная область характеризуется некоторыми особенностями (см. рис. 34, Б). Так, внешне она не столь выражена, и название "аурикулы" используется лишь постольку, поскольку оно употребляется в отношении всех анамний. С учетом того, что описательный термин "аурикулы" применяется для обозначения тех отделов мозжечка, которые связаны с вестибуло-латеральной областью мозга, полагают, что в мозжечке скатов идентичным отделом является так называемая вестибуло-латеральная доля, lobus vestibulolateralis. Она занимает вентролатеральное положение по отношению к телу мозжечка и разделена на непарную медиальную часть, pars medialis (гомолог одноименного отдела мозжечка акул), и парные латеральные
части, partes laterales (гомологичные верхнему листку аурикул акул).
Латеральная часть включает в себя дорсальный гранулярный гребень и латеральную гранулярную область. Они сходны по строению, но отличаются по связям: дорсальный гранулярный гребень "соседствует" с дорсальным ядром октаво-латеральной области, и его клетки формируют молекулярный слой ядра, в то время как латеральная гранулярная область аналогичным образом связана с медиальным ядром октаво-латеральной области продолговатого мозга.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|