- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 2 страница
К настоящему времени указанные коэффициенты вычислены практически для представителей всех групп современных позвоночных. Так, для круглоротых коэффициент аллометрии составил 0,23 - 0,41, хрящевых рыб - 0,60, костистых рыб - 0,49, амфибий - 0,56 - 0,60, рептилий - 0,66, птиц - 0,36, млекопитающих - 0,63 - 0,67. Определение этих величин позволяет получить информацию о характере эволюционных преобразований мозга позвоночных и оценить уровень развития их нервной системы.
Значения коэффициентов аллометрии, рассчитанные для группы и для отдельных видов, могут существенно различаться. В пределах каждой группы позвоночных встречаются особи с высокими и низкими индексами энцефализации (I), рассчитываемыми по формуле I =
| ky |
| k0 |
, где ky - коэффициент энцефализации данного животного, ky-коэффициент энцефализации, принятый за точку отсчета. Например, индексы энцефализации ряда представителей приматов, рассчитанные относительно индекса энцефализации примитивных насекомоядных, составили: лемур - 429-629, бабуин - 924, макака-резус - 892, капуцин - 1038, горилла - 668, орангутанг - 890, шимпанзе - 1131, человек - 2876.
С учетом этого обстоятельства в сравнительной нейрологии введено понятие "полигон энцефализации". Он представляет собой набор значений индексов энцефализации, определяемых отдельно для каждого вида и образующих некую геометрическую фигуру, ограниченную их крайними значениями. Такое построение позволило более тонко анализировать эволюционные изменения ЦНС в каждой конкретной группе позвоночных сравнительно с другими животными. Рассматривая полигоны
энцефализации для основных групп современных позвоночных (рис. 7), можно отметить ряд закономерностей. Так, полигон энцефализации птиц четко отделен от полигона энцефализации рептилий, который сам по себе включает самостоятельные части, соответствующие основным группам современных рептилий. Анализ индексов энцефализации представителей вымерших групп рептилий, проведенный по эндокранам (динозавры, птерозавры, маммалоподобные рептилии), показал, что и их полигоны энцефализации не достигают уровня млекопитающих. Полигон амфибий расположен отчасти в зоне полигона рептилий. Обширный полигон костистых рыб наглядно демонстрирует значительную эволюционную и адаптивную пластичность ЦНС у представителей многочисленных отрядов современных рыб, но и он, даже в своих высших значениях, нигде не пересекается с полигоном млекопитающих. Только хрящевые рыбы имеют индексы энцефализации, сравнимые с таковыми высших позвоночных - птиц и млекопитающих, что свидетельствует о высоком общем уровне организации их ЦНС.
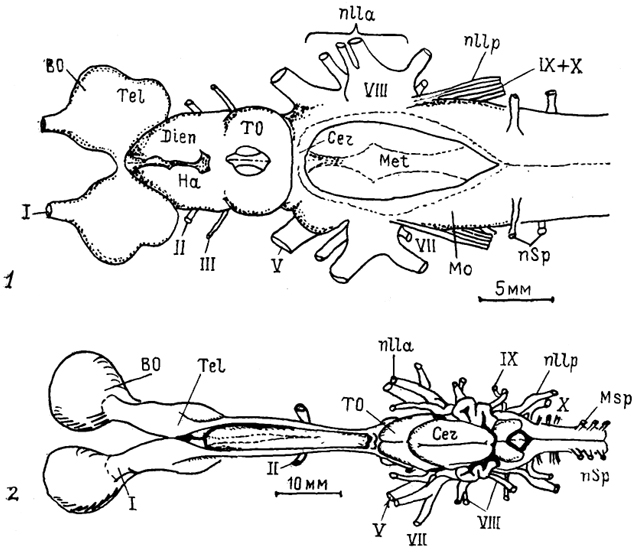
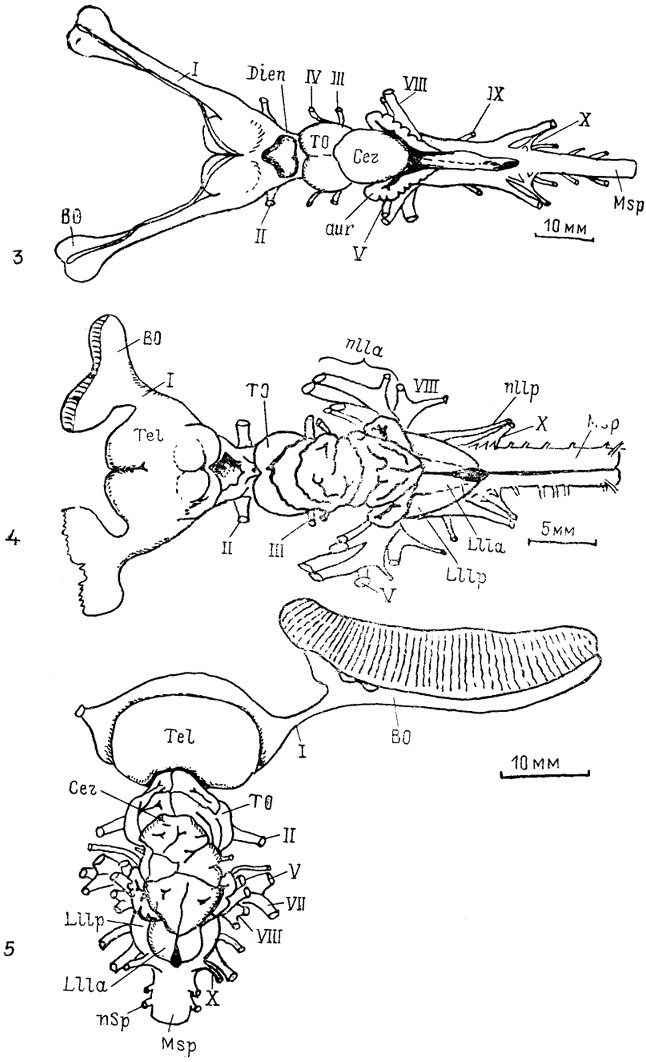
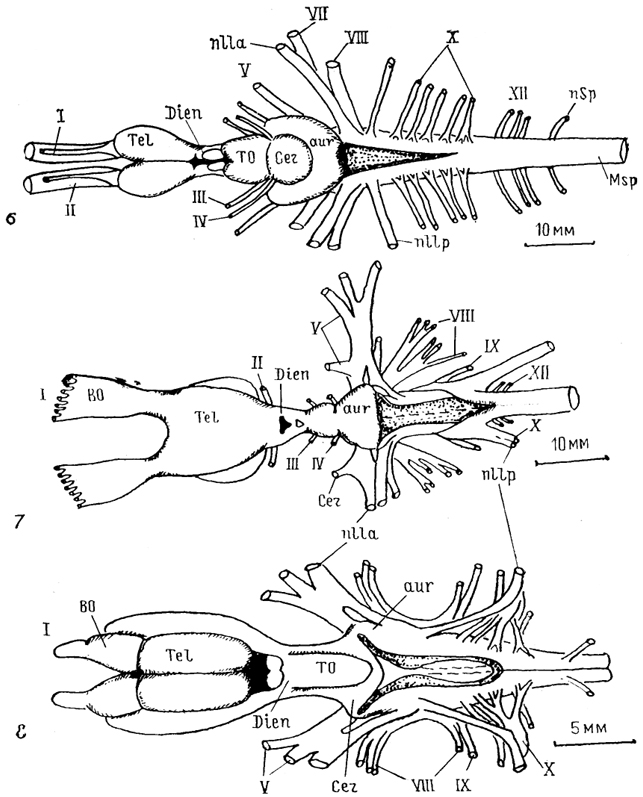
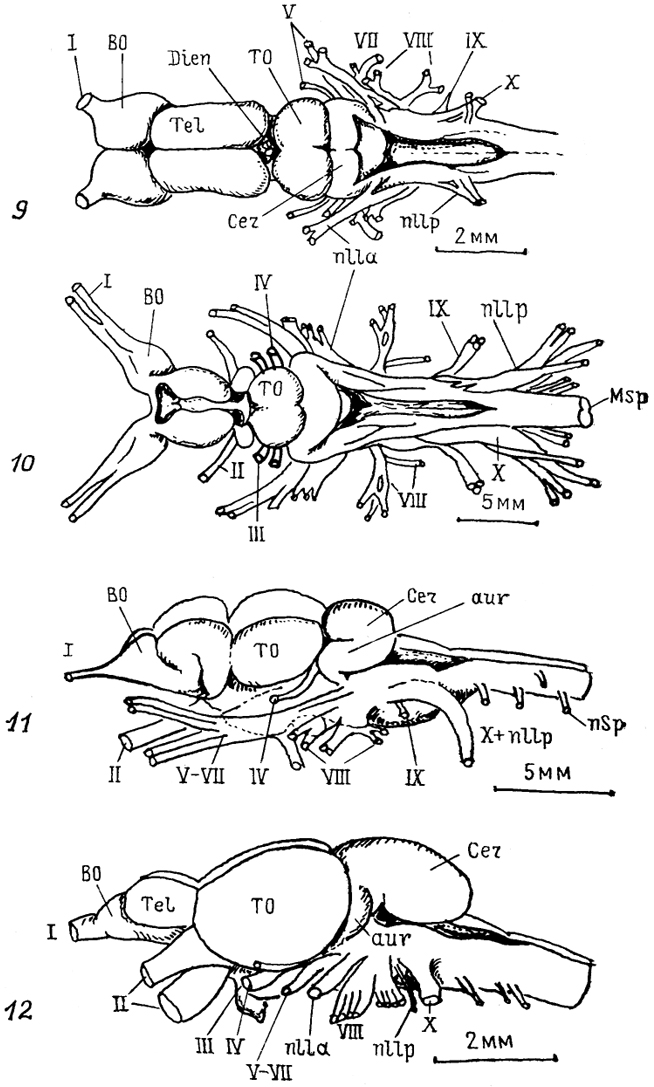
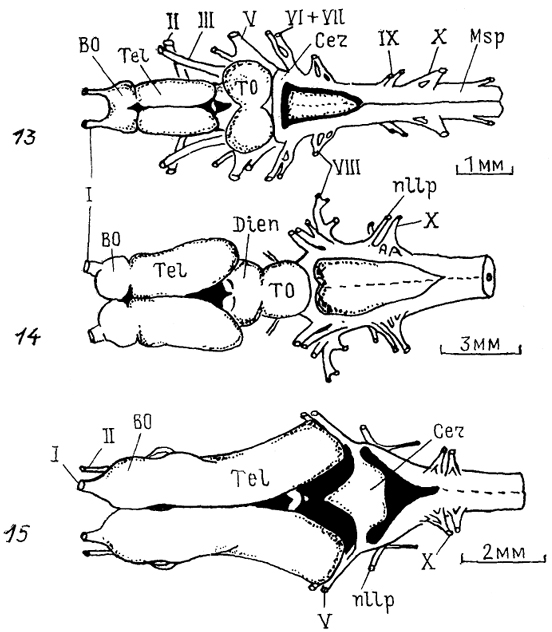
Рис. 5. Общий вид головного мозга представителей разных групп низших позвоночных (по: Northcutt, 1978, 1980, 1986; Northeutt, Puzdrowski, 1988)
1 - круглоротые (минога Ichthyomyzon unicuspis); 2-5 - хрящевые рыбы: химеры Hydrolagus colliei (2), акулы: Notorynchus maculatus (3), Mustelus canis (4), Spfiyrna tiburo (5); 6-8 - саркоптеригии: латимерия Latimeria chalumnae (6), двоякодышащие рыбы: Neoceratodus forsteri (7), Protopterus annecteus (8) ; 9 - 12 - лучеперые рыбы: многопер Polypterus palmas (9), хрящевой ганоид Scapharhynchus platorynchus (10), костный ганоид LepIdosteus osseus (11), костистая рыба Salmo gairdneri (12); 13 - 15 - амфибии: Rапа catesbeiana (13), Amblystoma tigrinum (14), Ichthyophis glutinosus (15); I-XII-черепномозговые нервы.
По мнению нейроморфологсв и нейрофизиологов, не всегда ясно адаптивное значение увеличения массы мозга, однако несомненно, что оно непосредственно не связано с явлениями
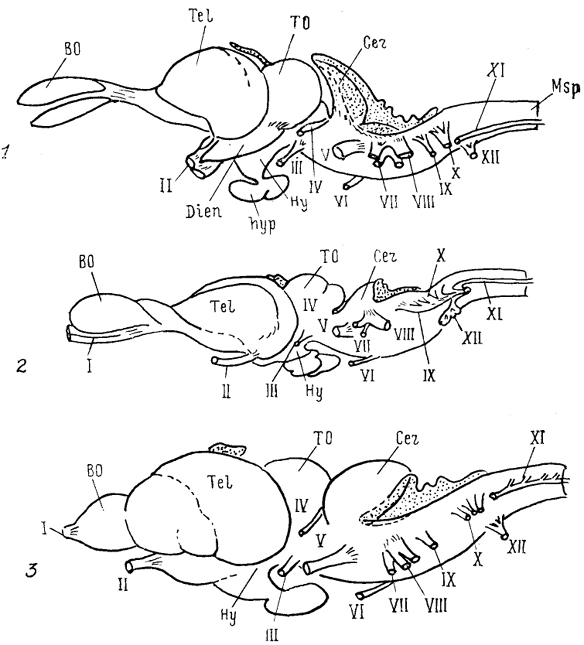
Рис. 6. Общий вид головного мозга рептилий (Bangma, 1983).
1 - ящерица Varanus exanthematicus, 2 - питон Pyton regius scripta elegans, 3 - черепаха Pseudemys.
пластичности и разнообразия функциональных связей в ЦНС. Таким образом, межвидовые и межгрупповые сравнения по массе, объему мозга или его отдельных частей, вычисление коэффициентов и индексов энцефализацни и аллометрии могут
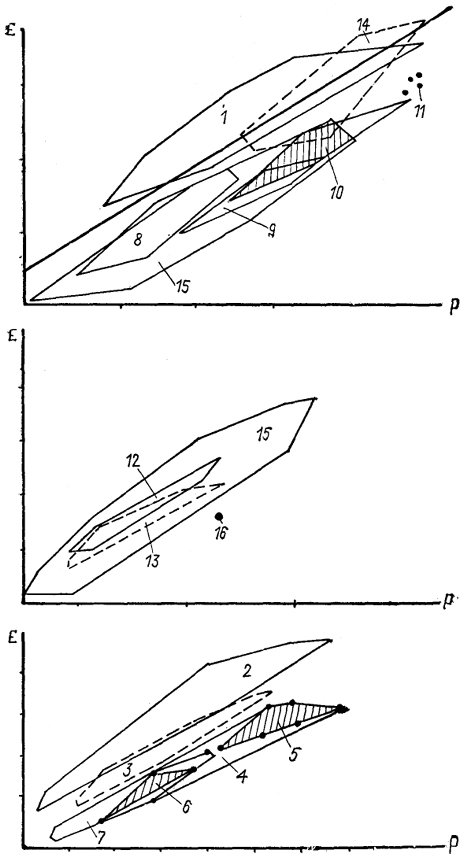
Рис. 7. Полигоны индексов энцефализации основных групп позвоночных (современных и вымерших) (Platel, 1984).
1 - птицы; 2 - современные и 3 - древние млекопитающие; 4-11 - рептилии: современные и древние формы (4), динозавры (5), древние маммалоподобные (6 , современные (7), ящерицы (8), змеи (9), черепахи (10), крокодилы(11); 12 - бесхвостые и 13 - хвостатые амфибии; 14 - хрящевые и 15 - костистые рыбы; 16 - минога; прямая линия - корреляционная кривая, рассчитанная для низших насекомоядных. Масштаб графиков не совпадает. По оси абсцисс - масса тела, по оси ординат - масса мозга
служить только первым шагом в анализе сложных процессов морфофункциональной эволюции нервной системы позвоночных.
* Часто в составе мозга низших позвоночных два слабо дифференцированных отдела - промежуточный и конечный мозг - объединяют названием передний мозг.
§ 2. СИСТЕМА ЖЕЛУДОЧКОВ МОЗГА
Представленные ниже описание и схема развития системы мозговых желудочков (рис. 8) получены при исследовании онтогенеза ЦНС у представителей млекопитающих. На стадии 20 сомитов (у человека приблизительно 24-й день развития) в головном отделе нервной трубки формируются три первичные мозговые полости (стадия трех мозговых пузырей), полость первичного переднего мозга (prosocoele), средняя мозговая полость (mesocoele) и полость мозгового ствола (rhombocoe1е). Остальная часть полости нервной трубки на этой стадии получила название syringocoele; на ее основе в дальнейшем образуется центральный спинномозговой канал.
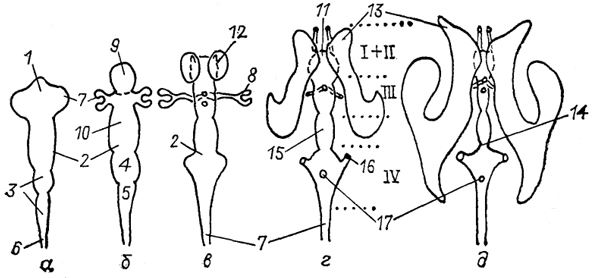
Рис. 8. Схема развития мозговых желудочков млекопитающих (McFarland е. а., 1969).
а-д - этапы развития мозговых желудочков на стадии трех (а) и пяти (б, в) мозговых пузырей, в пре- (г) и постнатальный (д)периоды. 1 - прозоцель и его производные: телоцель (9) и диоцель (10); 2 - мезоцель; 3 - ромбоцель и его производные: метацель (4) и миелоцель (5); 6 - сирингоцель и его производное - спинномозговой канал (7); 8 - глазной пузырь; 11, 12 - полость (11) и отверстия (12) Монро; 13 - боковые желудочки (I, II); 14 - перешеек; 15 - водопровод мозга; 16, 17 - мозговые отверстия Люшка (16) и Мажанди (17). Пунктиром обозначены границы I-IV мозговых желудочков.
Первичный конечный мозг развивается как вырост первого мозгового пузыря ростральнее оптической зоны (стадия пяти мозговых пузырей). Формирующаяся при этом борозда отделяет ростральную часть переднего мозга с его полостью (telocoele medialis) от каудальной части переднего мозга с его полостью (diocoele), из которой в дальнейшем образуется промежуточный мозг. Полость среднего мозгового пузыря на этой стадии еще превосходит по размерам полость переднего мозга. Полость ствола мозга разделяется на полости заднего (metacoele) и продолговатого (myelocoele) мозга.
На стадии 31 сомита в полости конечного мозга (telocoele) формируются билатеральные выпячивания, образующие боковые
желудочки, ventriculi laterales, которые отделяются от непарной части полости конечного мозга септальной бороздой. Полость непарной части конечного мозга в дальнейшем становится полостью Монро, которая соединяется с III мозговым желудочком (полостью промежуточного мозга) межжелудочковыми отверстиями, или отверстиями Монро. В то же время в развивающихся полушариях образуется еще одна борозда - ринальная, которая отделяет зачатки будущих обонятельных луковиц от остальной части полушарий.
В результате быстрого роста дорсального отдела таламуса латеральные стенки полости промежуточного мозга сближаются, образуя узкую щель - III мозговой желудочек, ventriculus tertius. Следует сказать, что данный процесс наблюдается лишь у высших млекопитающих, тогда как у всех остальных позвоночных объем III желудочка остается достаточно большим. Полость среднего мозга на этой стадии развития также еще большая и только впоследствии сужается, формируя водопровод мозга, aqueductus cerebri. В заднем мозговом пузыре образуются латеральные выросты, на концах которых формируются отверстия Люшка и Мажанди, играющие важную роль в циркуляции цереброспинальной жидкости. Отверстие Люшка, в отличие от отверстия Мажанди, есть у всех млекопитающих. У человека отверстие Мажанди на 5 - 6-м месяце эмбрионального развития открывается в полость мозговых оболочек.
Понимание процесса развития и изучение конфигурации мозговых желудочков в филогенезе позвоночных позволяют лучше ориентироваться при описании структур мозга (рис. 9). При этом важное значение приобретает проблема гомологии отдельных образований желудочковой системы у представителей разных групп. Даже в пределах одного класса, например млекопитающих, имеется значительная вариабельность в развитии тех или иных отделов.
Все мозговые желудочки и спинномозговой канал выстланы изнутри слоем специализированных эпендимных клеток, являющихся разновидностью глиальных элементов нервной ткани и происходящих из матричных клеток нервной трубки. Однако эпендимная выстилка мозговых желудочков неоднородна, в ней есть участки, имеющие атипическое строение - так называемые органы циркумвентрикулярной системы. Наиболее подробно она изучена у млекопитающих, где выделено несколько групп таких органов (рис. 10).
К первой группе органов циркумвентрикулярной системы принадлежат участки специализированной эпендимной выстилки, образующие сосудистые сплетения (PC III, PC IV), а также структуры парафиза (РА), дорсального и сосудистого мешков III мозгового желудочка.
Сосудистые сплетения находятся в полостях боковых и IV мозгового желудочков. В них происходит образование основного
объема цереброспинальной жидкости. Эпендимные клетки сплетений часто называют хориоидным эпителием. Они имеют кубовидную форму и на апикальной поверхности несут многочисленные микроворсинки. Плотные контакты между эпендимными клетками препятствуют прямому проникновению веществ из кровяного русла в полость желудочков.
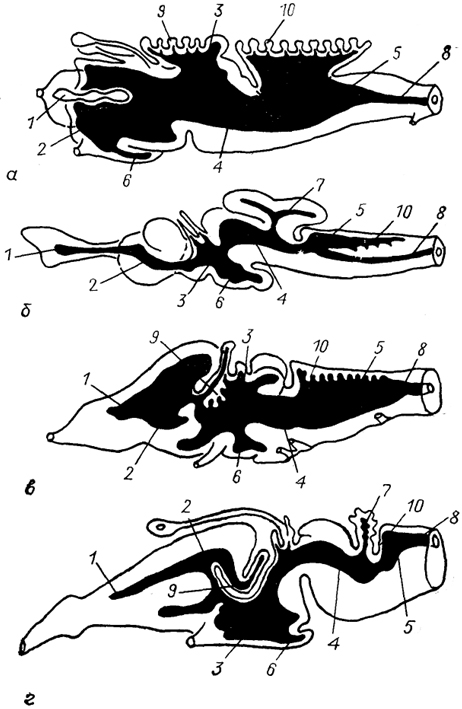
Рис. 9. Строение мозговых желудочков у ряда позвоночных (McFarland e. а., 1969).
а - минога, б - акула, в - лягушка, г - ящерица. 1 - обонятельные желудочки; 2-5 - латеральные (2), третий (3), среднемозговой (4), четвертый (5) желудочки; 6-8 - полость воронки гипоталамуса (6), мозжечка(7), спинного мозга(8); 9, 10 - сосудистые сплетения в III (9) и IV (10) мозговых желудочках.
Цереброспинальная жидкость (ЦСЖ) играет роль гидростатического
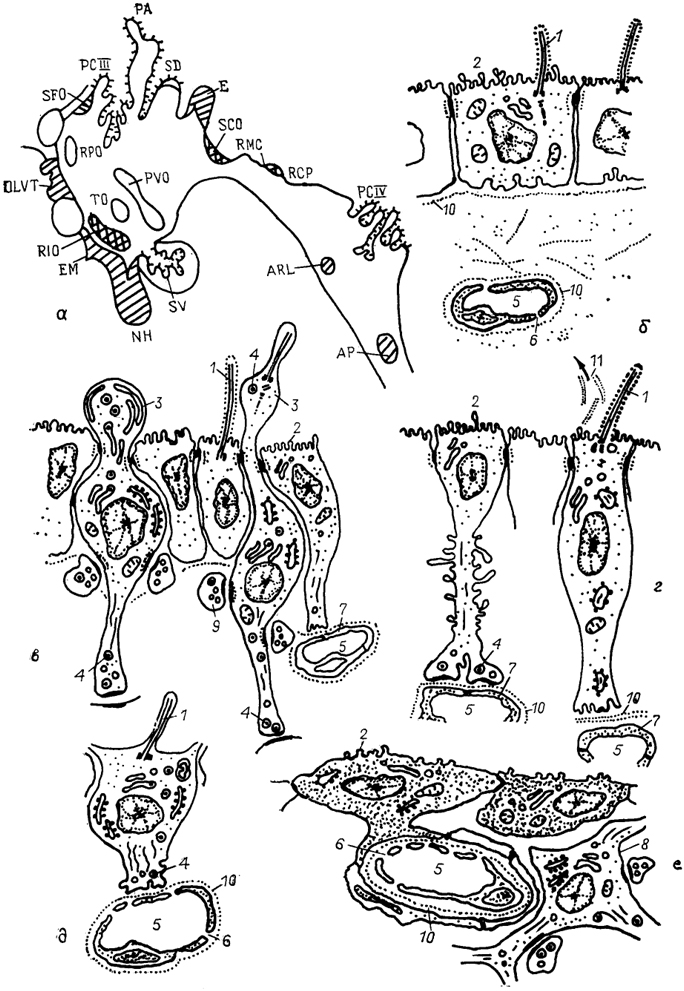
Рис. 10. Циркумвентрикулярная система мозга позвоночных (по: Oksche, 1973, 1988).
а - общий план строения; б-е - фрагменты ультраструктуры клеток эпендимы сосудистого сплетения (б), ликворконтактирующих нейронов преоптической зоны (в), танницитов субкомиссуального органа (г), пинеалоцитов эпифиза (д) и специализированной эпендимы area postrema (е). 1 - реснички; 2 - микроворсинки; 3 - выросты апикальной части клеток; 4 - секреторные гранулы и пузырьки; 5 - кровеносные капилляры; 6, 7 - эндотелий сосудов с порами (6) и без них(7); 8 - нейрон; 9 - синапсы; 10 - базальная мембрана; 11 - рейснеровские волокна. Объяснения в тексте.
буфера, заполняя пространства между мозговыми оболочками; поддерживает ионный баланс в мозговой ткани; является средой-переносчиком биологически активных веществ: нейромедиаторов, гормонов, нейросекретов; удаляет из нервной ткани продукты метаболизма, попаданию которых в кровь препятствует гемато-энцефалический барьер. Продуцирование ЦСЖ является активным процессом. Ее отток из желудочков осуществляется через полость ромбовидного мозга в подпаутинное пространство мозга через отверстия Люшка и Мажанди, а оттуда - в венозные синусы твердой мозговой оболочки.
Среди низших позвоночных степень развития сосудистых сплетений сильно варьирует. У круглоротых в переднем мозге они практически не развиты, что компенсируется значительным развитием сосудистого сплетения продолговатого мозга (рис. 9). У рыб выстилка сосудистых сплетений представлена эпендимными клетками самых разных типов: от кубовидных клеток, связанных с богатой сетью капилляров мягкой мозговой оболочки, до уплощенных клеток без ресничек и микроворсинок, фактически не участвующих в образовании ЦСЖ (например, в конечном мозге некоторых бесхвостых амфибий и костистых рыб).
Парафиз (РА) - наименее изученный орган циркумвентрикулярной системы. У некоторых хрящевых рыб, например акул и химер, парафиз имеет дольчатое строение, а цилиндрические эпендимные клетки, в отличие от аналогичных элементов сосудистых сплетений, лишены ресничек. У высших представителей хрящевых рыб (в том числе и высших акул) парафиз, по-видимому, дегенерирует. У костных рыб степень развития парафиза варьирует от весьма значительной (осетровые и ильные рыбы) до крайне слабой (представители некоторых семейств высших костистых рыб). Парафиз у амфибий выражен хорошо, у высших позвоночных - умеренно, а у млекопитающих во взрослом состоянии часто отсутствует.
Таким образом, парафиз хорошо развит у примитивных групп, как у рыбоподобных, так и у тетрапод, у высших же представителей этих групп он менее выражен. Происхождение парафиза неясно: одни авторы считают его самостоятельным отделом циркумвентрикулярнок системы, другие - производным сосудистых сплетений.
Сосудистый мешок (SV) найден практически у всех хрящевых и костных рыб, за исключением отдельных представителей,
где его отсутствие, по-видимому, вторично. У костистых рыб степень его развития обусловлена их образом жизни и средой обитания. Так, у пресноводных видов сосудистый мешок слабо-дифференцирован, тогда как у эвригалинных (например, Апguella, Hypomesus) он выражен очень хорошо. Однако такая корреляция не прослеживается у представителей других групп рыб: у ряда пресноводных хрящевых рыб, костных ганоидов и брахиоптеригий эта структура развита весьма значительно. Другая тенденция в эволюции сосудистого мешка заключается в его постепенной изоляции от III желудочка. Это направление развития четко прослеживается от низших рыб к высшим, а у некоторых костистых рыб (например, представителей отряда Sardinella) он вообще замкнут. Еще одна особенность строения сосудистого мешка связана с распределением участков специализированной эпендимы: у хрящевых и костных ганоидов она обычно находится только в зоне, прилежащей к нейрогипофизу, а у представителей высших отрядов костистых рыб образует всю выстилку сосудистого мешка. Неясно, имелся ли этот орган у предков позвоночных, а следовательно, остается открытым вопрос: первично или вторично его отсутствие у современных круглоротых?
Во второй группе органов циркумвентрикулярной системы выделяют две подгруппы. В первую входят субкомиссуральный орган (SCO), выросты эпендимной выстилки желудочка среднего мозга (RMC, RCP) и воронка гипоталамуса (RIO). В этих органах выстилка полости мозговых желудочков представлена участками специализированной эпендимы, клетки которой обладают ярко выраженной секреторной активностью. Эти клетки, получившие название "эпендимные таннициты", имеют как апикальные отростки, контактирующие с полостью желудочка", так и базальные отростки, заполненные секреторными гранулами и достигающие капилляров (рис. 10). Ко второй подгруппе относятся паравентрикулярный орган (PVO) и преоптический вырост (RPO), где наряду с танницитами в просвет желудочков выходят отростки специализированных нейросекреторных клеток преоптического и паравентрикулярного ядер гипоталамуса. Ликворконтактирующие нейроны этих образований секретируют в цереброспинальную жидкость биологически активные вещества - нейрогормоны, медиаторы, нейропептиды, которые затем через систему мозговых оболочек попадают в кровь.
Субкомиссуральный орган (SCO) обнаружен практически у всех представителей позвоночных и является, по-видимому, одним из самых филогенетически древних отделов циркумвен-трикулярной системы. У круглоротых SCO парный, в то время как у других позвоночных парными являются только его ростральные части. В некоторых группах млекопитающих (насекомоядные, китообразные, высшие приматы) с возрастом наблюдается
некоторая редукция SCO. Клетки этого органа отличаются ярко выраженной полярностью. Секрет выводится в ЦСЖ в виде фибриллярного материала - так называемых Рейснеровских волокон. Эти волокнистые структуры накапливаются в каудальной части центрального спинномозгового канала или в субарахноидальном пространстве в области задней комиссуры, откуда активно всасываются в кровь. Предполагают, что секрет SCO представляет собой гормон, участвующий в регуляции процессов роста и дифференцировки организма. SCO в онтогенезе позвоночных начинает функционировать очень рано и наиболее активен в первую половину эмбрионального развития.
Третья группа органов цирку мвентрикулярной системы представлена такими участками стенок мозговых желудочков, где эпендимная выстилка полностью или частично замещается на нервные или нейросекреторные клетки и имеется необычная система капилляров. К такого рода участкам относятся: субфорникальный орган (SFO), сосудистый орган конечной пластинки (OVLT), срединное возвышение (ЕМ), нейрогипофиз (NH), эпифиз (Е), расположенные в III желудочке, и зона латеральных выростов и area postrema (ARL, АР), расположенные в полости IV мозгового желудочка (рис. 10).
В субфорникальном органе (SFO) капилляры непосредственно контактируют с эпендимной выстилкой. У круглоротых и рыб SFO практически отсутствует, хотя участки, где крупные капилляры подходят к эпендиме, встречаются у многих костистых рыб в разных отделах мозга. Ряд исследователей связывает появление SFO в эволюции позвоночных с наземным образом жизни. Он хорошо развит у хвостатых амфибий, довольно слабо - у рептилий и птиц. У млекопитающих SFO сложно дифференцирован и включает в свой состав несколько типов нейронов, контактирующих с ЦСЖ и периваскулярным пространством капилляров. Полагают, что SFO принимает участие в регуляции водно-солевого обмена.
Area postrema (АР) - крупный орган циркумвентрикулярной системы, лежащий на дорсальной поверхности стенки желудочка, в каудальной части продолговатого мозга. АР представляет собой участок специализированного гематоэнцефалического барьера. Ее элементы выполняют нейросекреторную функцию, выделяя физиологически активные субстанции типа серотонина, энкефалина, холецистокинина и др., контролируют обмен натрия, регулируют кровообращение, дыхание, осморегуляцию, выделение и энергетический обмен. Эпендимные клетки АР лишены ресничек и связаны друг с другом плотными контактами. В паренхиме органа присутствуют группы глиальных клеток, нейронов и аксонных терминалей разного типа. Капилляры имеют многочисленные поры и окружены широким периваскулярным пространством, которое ограничено
плотной оболочкой из тел и отростков глиальных клеток (рис. 10).
Area postrema традиционно рассматривалась как область, присутствующая только в мозге млекопитающих и птиц. Однако современные исследования показали, что у многих представителей низших позвоночных она тоже имеется, и ее наличие или отсутствие не связано ни с филогенетическим положением, ни с образом жизни животного.
§ 3. УРОВНИ И ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
Для построения схем эволюционного развития ЦНС позвоночных необходим весь комплекс сведений об организации, нервной ткани: от анатомического описания отделов мозга до ультрамикроскопических, цитохимических и молекулярно-биологических данных. Все это дает возможность создать целостное представление об уровне организации и развития нервной: системы, оценить степень межвидовой и индивидуальной изменчивости структур нервных центров, уточнить "карты мозга".
Из множества нейрогистологических методов цитоархитектонический в наибольшей степени выявляет специфику мозга животных. Цитоархитектоника основана на анализе структурных различий в нервной ткани и использует для этого такие морфологические признаки, как величина и форма входящих в состав центра клеточных элементов, их число, плотность, наличие или отсутствие специализированных форм. Особое места отводится параметрам, связанным с пространственными, стереологическими характеристиками нервного центра: его объему, форме, характеру взаиморасположения клеток. Каждый участок мозга характеризуется определенным набором признаков. Значимость этих признаков различна: одни из них имеют принципиальное, первичное значение, рано появляются в онтогенезе и могут быть положены в основу разделения мозга на крупные области или зоны (например, при выделении формаций древней, старой и новой коры), другие, более частные, имеют вторичное значение, в онтогенезе появляются позднее и служат для выделения в составе основных подразделений более мелких структур (например, отделы коры, ее поля и подполя).
Кроме структурных признаков при морфологическом разделении мозга необходимо учитывать данные о его гистохимической природе, хемоархитектонике, системе афферентных и эфферентных связей (ходологии). Вместе с тем при проведении сравнительно-морфологического исследования и составлении цитоархитектонических карт мозга необходимо учитывать фактор внутри и межвидовой изменчивости, которая может достигать больших значений. Так, величины индивидуальной, внутривидовой, изменчивости в новой коре достигают 10 - 15%,
что вызывает определенные трудности при сопоставлении карт мозга.
В настоящее время, благодаря методам автоматизированного анализа, стало возможным получать количественную информацию о стуктуре мозга на качественно новом уровне. Особенно перспективен такой подход при изучении пространственных, стереологических, характеристик нервных центров.
Нейронный уровень
Наиболее ярко эволюционные и адаптивные изменения нервных центров проявляются в структуре дендритных ветвлений нейронов. Поэтому вполне естественны попытки нейроморфологов создать классификацию нейронов, основанную на этом признаке. Так, Э. Рамон-Молинер предложил разделить все нейроны на три основных класса: изодендритные, аллодендритные и идиодендритные.
Класс изодендритных (неспециализированных) нейронов характеризуется относительно слабым развитием дендритнога поля: отростки не многочисленны, ветвятся слабо. В зависимости от их ориентации среди изодендритных нейронов выделены лептодендритные, лофодендритные и радиальные.
Класс аллодендритных (специализированных) нейронов отличается значительным усложнением системы дендритов. Это выражается в определенной ориентации отростков, появлении: специфических форм дендритного поля. К аллодендритным нейронам относятся, например, пирамидные нейроны коры и. многие нейроны таламических ядер.
Класс идиодендритных (высокоспециализированных) нейронов объединяет большую группу клеток различных отделок ЦНС, отличающихся высокодифференцированной системой дендритного ветвления. Морфологически это выражается в резком увеличении разветвленности отростков, в появлении узлов ветвления и системы возвратных коллатералей.
На основе данной классификации была составлена схема возможных преобразований нейронной структуры мозга позвоночных (рис. 11). Полагают, что для мозга гипотетического предка позвоночных характерны нейроны двух основных типов: лепто- и лофодендритные, расположенные в основном в перивентрикулярных областях мозга. В эволюции, вероятно, происходило изменение локализации этих двух семейств нейронов, что привело к появлению перивентрикулярных и субповерхностных слоев мозга. Максимального выражения этот процесс достигает у млекопитающих, где он проявляется в двух основных тенденциях усложнения нейронной структуры мозга - теленцефалической и стволовой. Теленцефалическая тенденция (описанная еще С. Рамон-и-Кахалем) проявляется в появлении большого количества специализированных типов нейронов
с определенно ориентированными дендритными стволами (например, гранулярные нейроны гиппокампа, пирамидные нейроны новой коры, клетки Пуркинье мозжечка, нейроны сетчатки и обонятельной луковицы). Вторая линия развития - стволовая - связана с преобразованием изодендритных нейронов ствола в систему высокоспециализированных, идиодендритных, клеток (например, "кисточковые" нейроны специфических центров ствола и промежуточного мозга). Следует отметить, что при этом значительная часть изодендригных клеток сохраняется в области осевой ретикулярной формации.
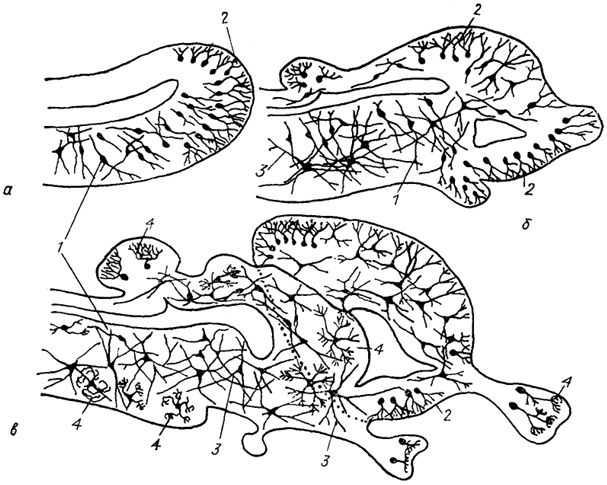
Рис. 11. Схема возможной эволюции нейронной структуры мозга позвоночных (Ramon-Moliner, 1975).
а - мозг гипотетического предка позвоночных; б, в - современных позвоночных низших (б) и высших (в). 1-4 - различные типы нейронов: лептодендритные (1), лофодендритные(2), аллодендритные (3), идиодендритные (4).
Классификация Рамон-Молинера в настоящее время широко используется нейроморфологами для изучения эволюционных преобразований структуры мозга. Более того, подобный подход позволяет провести теоретический анализ межнейронных взаимодействий в нервных центрах. Так, при равенстве линейных параметров дендритов (длина, диаметр) объем, занимаемый собственно дендритами, по отношению к объему дендритного поля нейрона будет наименьшим для изодендритных клеток, большим для аллодендритных и максимальным для идиодендритных клеток. Эта зависимость имеет важное значение
для понимания организации взаимоотношений нейронов с афферентными волокнами. Так, в нервном центре, образованном изодендритными клетками, будет больше контактов между
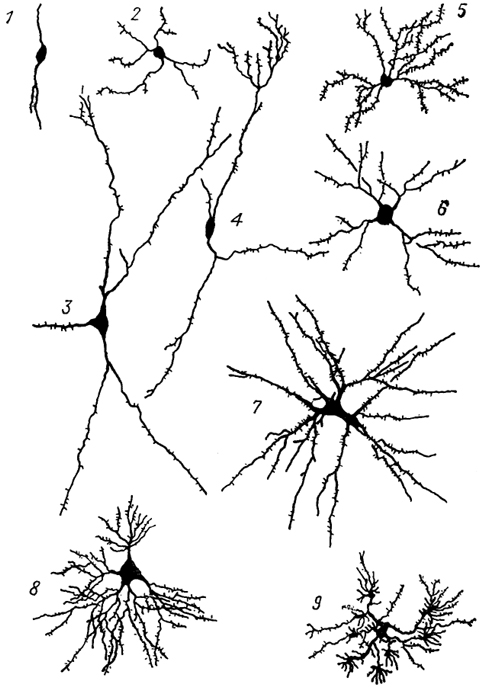
Рис. 12. Различные виды длинноаксонных нейронов подкорковых центров переднего мозга собаки (Леонтович, 1978).
1-3 - редковетвистые нейроны: нейробластоформный (1), короткодендритный (2), ретикулярный (3); 4-9 - густоветвистые нейроны: штамбовый (4), древовидные: густо-шипиковый (5) и редкошипиковый (6), мультиполярный гигантский (7), кустовидный (8) и кисточковый (9).
дендритами данной клетки и различными аксонами, входящими в этот центр, т.е. связи будут неспецифическими, случайными, тогда как в нейронных популяциях аллодендритного или идиодендритного типов будет больше контактов, образованных дендритами данной клетки с определенными аксонами, т.е. связи будут более специфическими.
Классификация Т.А. Леонтович акцентирует внимание на количественных характеристиках дендритной системы нейронов.
Рис. 13. Схема распределения элементов редковетвистой и густоветвистой нейронных систем в структурах головного мозга (Леонтович, 1978).
1, 2 - нейронные системы мозга: густоветвистая (1) и редковетвистая (2); 3 - ретикулярные клетки в структурах густоветвистой системы; 4 - густоветвистые (мультиполярные гигантские) клетки в структурах редковетвистой системы.
Все нейроны разделяют на две категории: длинноаксонные и короткоаксонные, в каждой из которых описано несколько классов клеток. Длинноаксонные нейроны разделяются на редко- и густоветвистые. Редковетвистые включают три класса клеток: нейробластоформные, короткодендритные и ретикулярные, а густоветвистые - пять классов: мультиполярные гигантские, штамбовые, древовидные, кустовидные и кисточковые (рис. 12). Каждый класс нейронов объединяет несколько разновидностей клеток и характеризуется определенным набором количественных признаков. Классификация Т.А. Леонтович разработана на мозге млекопитающих, однако она с успехом применяется для изучения нейронной структуры мозга позвоночных других групп, поскольку основана на универсальных, единых для всех нейронов, принципах ветвления дендритов и аксонов.
Исследование нейронной структуры мозга позвоночных различных рангов позволило выдвинуть и обосновать ряд общих принципов ее развития.
В мозге позвоночных не происходит "отрицания" старых форм и типов клеток, отделов. и даже целых систем мозга. В процессе эволюции они дополняются новыми образованиями, преобразуются или сохраняют принципы своего строения неизменными (принцип дополнительности). Это приводит к значительной гетерогенности нейронного состава многих отделов мозга, где наряду с эволюционно новыми, высокодифференцированными элементами часто сохраняются элементы, свойственные представителям более древних групп позвоночных. Об этом, в частности, свидетельствует факт "проникновения" ретикулярных редковетвистых нейронов - одной из наиболее филогенетически древних форм нейронов, характерной
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|