- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 3 страница
для стволовых отделов мозга позвоночных - во все вышележащие отделы, включая конечный мозг (рис. 13).
Большое количество нейронов совмещает в себе черты строения разных клеточных типов, причем у высших позвоночных число таких "переходных" клеточных форм значительно возрастает (принцип структурной переходности). Значение их в функционировании нервных центров достаточно велико - благодаря сочетанию признаков, характерных для разных типов нейронов, они могут выполнять более широкие функциональные задачи, нежели крайние по форме нейронные типы.
На цитоархитектоническом уровне принцип структурной переходности проявляется в присутствии в ряде нервных центров (особенно в коре больших полушарий) межуточных формаций, соединяющих разные по строению и происхождению отделы. Данный принцип, в частности, свидетельствует о многоэтапности и неравномерности развития нервной системы в эволюции позвоночных.
Нейроны одного вида различаются между собой рядом признаков клеточной структуры, вследствие чего возможно определить лишь границы типовой формы нейронов (принцип полиморфизма). Так, даже у одного нейрона дендритные и аксонные ветви отличаются друг от друга по многим признакам, что обусловливает многообразие переходных форм нейронов. В процессе эволюции полиморфизм нейронной структуры мозга нарастает; причем это выражается, в частности, в усложнений ветвления отростков.
Синаптический уровень
В нервной системе выделяются три основные группы синаптических контактов: электротонические, химические и смешанные, электро-химические. Они обнаружены практически во всех отделах ЦНС позвоночных разных групп и различаются между собой - в гомологичных центрах или нейронах - по форме синаптичеокого бутона, размеру, количеству и характеру распределения синапсов на теле и отростках нейронов.
Электротонические синапсы представляют собой особую разновидность щелевого межклеточного контакта, приспособленного для прямой передачи нервного электрического импульса с одной клетки на другую. Как и в других видах синапсов, здесь выделяют пре- и постоинаптическую части и синаптическую щель. Однако есть и специфические черты организаций: отсутствие субмембранных специализаций пре- и постсинаптических мембран, узкая, 2-3 нм, синаптическая щель, наличие в мембранах особых белковых комплексов - коннексонов, контактирующих в синаптической щели друг с другом и обеспечивающих проведение импульса, отсутствие, как правило, в пресинапсе синаптических пузырьков. Такая структура
контакта обусловливает его быстродействие, надежность в работе, практическую неутомляемость, возможность проведения импульса в обоих направлениях. В отношении последней характеристики необходимо подчеркнуть, что в ряде нейронных систем мозга позвоночных (например, в системе мюллеровских и маутнеровских клеток круглоротых и рыб) импульсы передаются в строго определенном направлении - от "командных" мюллеровских и маутнеровских нейронов. Электротонические синапсы обнаружены во многих отделах нервной системы у всех изученных представителей позвоночных.
Таблица 1. Механизмы передачи в гомологичных синапсах ЦНС (Шаповалов, Ширяев, 1987)
| Класс позвоночных | Нейроны, образующие синапс | ||
| первичный афферент-мотонейрон | ретикулоспинальный нейрон-мотонейрон | вестибулярный афферент-ядро Дейтерса | |
| Круглоротые: минога | Смешанный | Смешанный | Смешанный |
| Амфибии: лягушка | -"- | Химический или смешанный | Химический или смешанный |
| Млекопитающие: крыса кошка | Химический -"- | Химический -"- | -"- Химический |
Химические синапсы характеризуются наличием в пресинапсе синаптических пузырьков, развитой системой пре- и постсинаптической субмембранных специализаций ("а основании этого выделяют две разновидности синапсов - симметричные и асимметричные), достаточно широкой синаптической щелью (от 20 до 50 нм). Показано, что любая часть нейрона - тело, аксон, дендриты - может образовывать синапсы. Наибольший интерес представляют аксо-аксонные и дендро-дендритные контакты, поскольку они имеют важное значение для обеспечения процессов авторегуляции деятельности нейронов (при взаимодействии аксонных коллатералей с аксонным холмиком собственного нейрона) к для осуществления тормозных взаимодействий в нейронных цепях. Очень часто соединения подобных типов являются контактами реципрокными, т. е. имеющими по две пре- и постсинаптические зоны, что обеспечивает двустороннее проведение импульса. Встречаются и так называемые смешанные, электро-химические, контакты.
Вопрос о происхождении различных типов синапсов в гомологичных нервных центрах мозга позвоночных разного филогенетического уровня не решен. Одни исследователи рассматривают электротонические синапсы как первичные в эволюции нервной системы и указывают на существование в филогенезе процесса замещения электротонических контактов химическими

Рис. 14. Синаптические комплексы различного вида в структурах нервной системы млекопитающих.
а-е - дивергентные комплексы в структурах моста (а), кохлеарном ядре (б), вентробазальном таламусе (в), коре мозжечка (г), желатинозной субстанции спинного мозга (д), клиновидном ядре продолговатого мозга (е); ж, з - конвергентные комплексы на нейронах таламуса (ж) и мотонейронах спинного мозга (з). 1 - аксоны, 2 - дендриты, 3 - дендритные шипики. Стрелки - тормозные воздействия.
(табл. 1). Другие считают, что как электротонические, так и химические синапсы возникли в эволюции нервной системы очень рано и имеют широкое распространение в нервных центрах мозга позвоночных всех филогенетических групп. Преобладание в ряде нервных центров того или иного типа синаптической
передачи определяется функциональными задачами данного центра, а не его происхождением. Эта точка зрения базируется на фактах обнаружения типичных химических и электротонических синапсов в нервной системе низших многоклеточных животных с одной стороны, и в высших интегративных центрах конечного мозга млекопитающих и птиц, с другой. Более того, показано, ч го синапсы, образуемые одним аксоном на разных постсинаптических элементах, могут работать с использованием и химического, и электротонического механизмов.
Связи между нейронами могут осуществляться как одиночными синапсами различного вида, так и сложными комплексами типа "гломерул" (рис. 14). Синаптические комплексы построены по двум основным принципам: конвергенции и дивергенции. В центре конвергентного синаптического комплекса находится один постсинаптический элемент - тело или отросток нейрона - на котором оканчивается несколько пресинапсов различного происхождения и типа. В дивергентном комплексе отношения обратные: одна пресинаптическая часть контакта - чаще всего окончание или расширение аксона - оказывает влияние на несколько постсинаптических элементов.
В последние годы во многих отделах нервной системы позвоночных обнаружены так называемые модуляторные контакты. Нейроактивная субстанция в них заключена, как правило, в крупные секреторные гранулы. Путем экзоцитоза она выделяется в межклеточные пространства и, достигая рецепторов постсинаптических клеток, воздействует не на ионную проницаемость мембраны (как в типичном химическом синапсе), а на синтетические процессы в клетках-мишенях, изменяя, в частности, уровень синтеза цАМФ, активность мембранной АТФазы или протеинкиназ. Имеется еще ряд существенных отличий в организации и функционировании синаптических и модуляторных контактов в нервной системе позвоночных (табл. 2).
Важно отметить, что термины "нейромедиатор", "нейромодулятор", "нейрогормон" отражают скорее механизм взаимодействия этих соединений с клетками-мишенями, чем их химическую природу. Одно и то же вещество в разных отделах нервной системы организма может выступать как в роли медиатора, так и нейромодулятора или нейрогормона. Многие нейропептиды, например энкефалины и эндорфины, ранее рассматриваемые только как нейрогормоны, выделяются аксонными терминалями и выступают в роли типичных нейромедиаторов. И наоборот, отдельные биогенные амины (адреналин, дофамин, серотонин) не только действуют через межнейронные синапсы, но могут также выделяться нейросекреторньши клетками гипоталамических ядер или органов циркумвентрикулярной системы в кровь и цереброспинальную жидкость, распространяясь
затем в организме на значительные расстояния и действуя как типичные нейрогормоны.
Таблица 2. Различия между контактами в ДНС позвоночных (по: Vizi, 1984)
| Параметры | Контакты | ||
| синаптические | модуляторные | нейросекреторные* | |
| Тип связи | Один на один | Один на много | Один на много |
| Время действия | Короткое, мс | Длительное, с | Длительное, мин и ч |
| Механизм действия | На ионную проницаемость мембран | Без прямого действия на ионную проницаемость мембран | |
| Характер действия | Фазный | Тонический | Тонический |
| Место действия | Пре- и пост-синапс | Пре- и пост-синапс | Пре- и пост-синапс, клетки-мишени, органы |
| Место выведения | Синаптическая щель (5 - 50 нм) | Межклеточные пространства, мкм, мм | Межклеточные пространства, кровь, спинномозговая жидкость |
| Активность соединений | Низкая | Средняя | Чрезвычайно высокая |
| Специфичность соединений | Высокая | Высокая | Высокая |
| Скорость синтеза | Средняя | Средняя | Низкая |
| Молекулярный вес | Небольшой | Небольшой | Небольшой и средний |
| Концентрация молекул | Высокая(10-4 - 10 3M) | Средняя(10 6 - 10-5M) | Низкая(10-9 - 10-12M) |
*Нейросекреторные окончания образованы отростками специализированных клеток, составляющих важный интегрирующий компонент нервной системы. Нейросекреторные клетки - филогенетически древние элементы нервной системы, сочетающие в себе свойства нервных и секреторных клеток. Они синтезируют и выделяют целый комплекс биологически активных веществ - нейрогормонов. В настоящее время описано несколько десятков соединений, в основном пептидной природы, выполняющих эту важную функцию.
С помощью биохимических, авторадиографических и иммуноцитохимических методов в ЦНС выявлено несколько десятков таких соединений. При этом нейроны могут синтезировать и выделять в своих терминалях целый комплекс посредников (до 12!). В исследованиях на культуре нейронов показано, что клетки синтезируют несколько физиологически активных межнейронных посредников, сочетание которых в разных нейронах варьирует. Так, в 217 изученных нейронах было отмечено 20 комбинаций из 4 идентифицированных веществ, причем при одинаковом наборе посредников их количественное соотношение оказалось разным.
Способ накопления веществ в одном синапсе может быть различным: в одних и тех же синаптических пузырьках, в разных
популяциях пузырьков и секреторных гранул, в цитоплазме аксонной терминалы и пузырьках. В связи с этим наиболее сложным в проблеме межнейронных взаимодействий является исследование механизма действия различных субстанций, выделяемых одной аксонной терминалью. Существует два основных варианта действия медиаторов: дивергентный и независимый, при котором каждый выделяемый из терминали медиатор действует на определенную популяцию клеток-мишеней; и конвергентный и зависимый, при котором один медиатор оказывает определенное влияние на взаимодействие постсинаптической клетки с другим медиатором на разных этапах синаптической передачи.
Таким образом, краткий обзор строения и функций синаптических контактов в ЦНС позвоночных показал, что на фоне относительного единообразия структурной организации синапсов и консервативности химических соединений, используемых для передачи нервного импульса, в нервных центрах имеется возможность их эволюционного развития и усложнения путем комбинации различных типов межнейронных соединений (принцип полиморфизма) и использования различных посредников - медиаторов, модуляторов, нейрогормонов - в организации нейронных цепей.
Модульный уровень
Идея о том, что нервные центры состоят не из отдельных цепочек нейронов, работающих независимо друг от друга, а организованы в блоки или модули, объединяющие от нескольких клеток до нескольких тысяч нейронов, которые функционируют как единое целое, была выдвинута С. Рамон-и-Кахалем еще в начале XX в.
В ЦНС позвоночных модули были впервые обнаружены и описаны в коре мозжечка с его упорядоченным расположением •слоев и клеточных элементов. Позднее на тангенциальных срезах соматосенсорной коры мозга мыши были выявлены кольцеобразные скопления нейронов, названные "бочонками" (англ, barrels). Оказалось, что число и пространственное расположение этих модулей точно соответствует числу и расположению чувствительных вибрисс на морде животного. Это явилось одним из первых доказательств того, что модуль является структурно-функциональной единицей нервного центра. В дальнейшем модули были обнаружены в разных отделах ЦНС у многих позвоночных. Пространственная организация модулей может быть различной: от плоских дисков, характерных для нейропиля спинного мозга, до сложнейших пространственных комплексов- "бочонков" в новой коре и стриосом в базальных ядрах конечного мозга. Однако независимо от конкретного вида модуля, он представляет собой популяцию нейронов, способных
к возбуждению или торможению относительно независимо от подобных процессов в соседних группах нейронов. В состав любого модуля входят три группы клеток: нейроны, принимающие афферентные воздействия, эфферентные клетки, а также нейроны, осуществляющие локальные связи. Межнейронные контакты внутри модуля осуществляются системой "микромодулей", структурной основой которых могут быть пучки дендритов и аксонов.
Размеры и количество модулей в разных отделах ЦНС и у разных животных колеблются в широких пределах. Так, в новой коре млекопитающих насчитывается от 0,6 до 2-3 млн. объединений нейронов, в составе которых находится от 3 до 10 тыс. клеток. Размеры модуля определяются объемом ветвления афферентов, входящих в данный модуль. Элементарные модули могут объединяться в более сложные комплексы, содержащие до нескольких десятков модулей. Морфологической основой такого рода объединения могут служить аксонные коллатерали нейронов модуля, распространяющиеся на большие расстояния (в новой коре от 3 до 8 мм).
Таким образом, центральная нервная система всех позвоночных имеет единый план строения, основанный на общих принципах формирования и развития нервных центров всех уровней организации.
ГЛАВА 2
СПИННОЙ МОЗГ
Спинной мозг, Medulla spinalis, является центром, воспринимающим разнообразную соматическую информацию из внешней и внутренней среды и передающим ее в вышележащие отделы ЦНС. В нем сосредоточены моторные (эфферентные) центры, управляющие рефлекторной деятельностью поперечнополосатых мышц туловища и конечностей, и ассоциативные центры симпатического и парасимпатического отделов вегетативной нервной системы.
§ 1. ОБЩИЕ ПРЕДСТАВЛЕНИЯ ОБ ОРГАНИЗАЦИИ СПИННОГО МОЗГА ПОЗВОНОЧНЫХ
Анатомически спинной мозг разделяется на отделы, количество сегментов в которых соответствует числу позвонков у представителей данной группы позвоночных. Так, у птиц выделяют 12 шейных, 8 грудных, 12 поясничных и 6 копчиковых сегментов; у приматов - 8 шейных, 12 грудных, по 5 поясничных и крестцовых и 2-3 копчиковых, а у некоторых рептилий (змей) число спинномозговых сегментов может достигать 500.
У большинства позвоночных (рыбы, хвостатые амфибии, рептилии, птицы) спинной мозг занимает весь позвоночный канал, тогда как у бесхвостых амфибий и млекопитающих он занимает только часть канала из-за неравномерности роста спинного мозга и позвоночника в период эмбрионального развития. Так, у большинства млекопитающих спинной мозг оканчивается на уровне поясничных позвонков, однако у примитивных млекопитающих (нааример, ехидна) конец спинного мозга находится на уровне грудного отдела позвоночника. Наиболее короткий спинной мозг отмечен у южноамериканской лягушки Pipa pipa - он оканчивается на уровне 3-го позвонка. При этом для всех позвоночных выполняется одно правило: спинномозговые нервы каждого сегмента выходят из позвоночного канала через межпозвонковые отверстия на уровне
"своих" позвонков, в связи с чем в каудальной части позвоночного канала формируется особая анатомическая структура - пучок спинномозговых корешков, конский хвост.
На уровне каждого сегмента с обеих сторон спинного мозга отходит пара спинно-мозговых корешков: задний (дорсальный) и передний (вентральный) (рис. 15). У круглоротых дорсальные и вентральные корешки спинного мозга отходят асимметрично, тогда как у всех других позвоночных - симметрично.
В районе межпозвонковых отверстий спинномозговые корешки у большинства позвоночных (за исключением бесчерепных, миног и некоторых рыб) соединяются в единый спинномозговой нерв.
В составе задних корешков спинного мозга находятся спинномозговые ганглии (каждый корешок имеет по одному ганглию), содержащие чувствительные (афферентные) нейроны, которые воcпринимают
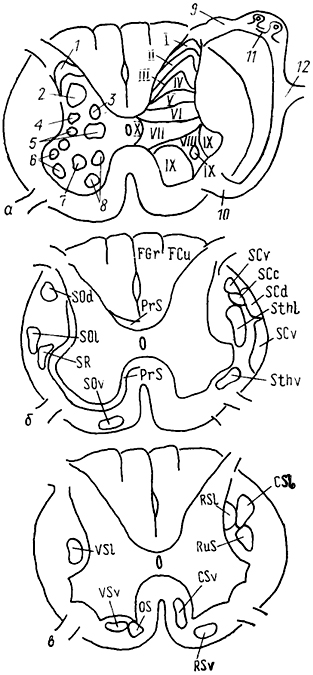
Рис. 15. Схема организации серого вещества (а) и топография восходящих (б) и нисходящих (в) трактов спинного мозга.
а: 1-Х - пластины серого вещества по классификации Б. Рекседа; 1-4 - желатинозная субстанция (1), собственное ядро (2), ядро Кларка (3), ретикулярное (4) ядро задних рогов; 5 - промежуточная зона, 6-8 - моторные ядра передних рогов: латеральные (6) , центральные (7), медиальные (8); 9, 10 - задний (9) и передний (10) спинномозговые корешки; 11 - спинальный ганглий, 12 - спинномозговой нерв.
б: восходящие тракты - спинно-мозжечковые: дорсальный SCd, вентральный SCv, ростральный SCr и кунеальный SCc; спинно-таламические: латеральный SThI и вентральный SThv. спинно-ретикуло-таламический SRT, спинно-тектальный ST; спинно-оливарные: дорсальный SOd, латеральный SOl и вентральный SOv, спинно-ретикулярный SR, нежный FGr и клиновидный FCu пучки; проприоспинальные PrS (собственные) пути спинного мозга (восходящие и нисходящие).
нисходящие тракты - кортико-спинальные: латеральный CSl и вентральный CSv, руброспинальный RuS; вестибуло-спинальные: латеральный VSI и передний VSv; ретикуло-спинальные: латеральный RSl и вентральный RSv; оливо-спинальный OS.
разнообразную информацию из кожи, опорно-двигательного аппарата, внутренних органов. Спинальные ганглии развиваются из элементов ганглионарной пластинки. Принципиальным является тот факт, что у ряда низших позвоночных (взрослые круглоротые, личинки рыб, амфибий) чувствительные нервные клетки располагаются не только в спинальных ганглиях, но и внутри спинного мозга. Эти клетки, получившие название "клетки Рогон-Берда", или дорсальные чувствительные клетки, находятся в дорсолатеральной части спинного мозга и представляют собой крупные (8-20 мкм) нейроны. От их тел отходят по два отростка, которые направляются вдоль спинного мозга, образуя синапсы (в основном
Таблица 3. Классификация афферентных волокон спинного мозга
| Тип | Диаметр, мкм | Миелинизация | Функциональная характеристика | |
| по Гассеру | по Ллойду | |||
| Аα | Ia, Ib | 12 - 20 | + | Проведение возбуждения от: первичных рецепторов мышц и сухожилий; |
| Аβ | II | 6 - 12 | + | от вторичных рецепторов мышц и рецепторов кожи; |
| Аδ | III | 4 - 6 | + | от рецепторов кожи и внутренних органов; |
| С | IV | 1,5 - 4 | - | от рецепторов сосудов, кожи, мышц |
электротонические) на телах и отростках спинальных интернейронов. Последние в свою очередь контактируют с мотонейронами, таким образом формируя первичную рефлекторную дугу сомато-моторного рефлекса. Электрофизиологические исследования показали, что клетки Рогон-Берда и замещающие их на более поздних этапах эмбриогенеза так называемые дорсальные чувствительные клетки связаны с механорецепцией,. тогда как чувствительные клетки в спинномозговых ганглиях воспринимают болевые и температурные стимулы. На теле клеток Рогон-Берда обнаружены многочисленные синаптические контакты неизвестного происхождения. Полагают, что они модулируют выделение медиатора (вещество Р) из клеток. У высших позвоночных на телах чувствительных нейронов в; спинномозговых ганглиях обнаружены синаптические контакты, которые образованы аксонами спинальных нейронов, приходящими по обоим корешкам.
Структура спинномозгового ганглия достаточно сложная. Морфологически в нем выделяются три основные группы нейронов: крупные и мелкие псевдоуниполярные и атипичные. Последние осуществляют внутриганглионарные связи. Более многочисленные
(85%) псевдоуниполярные нейроны являются сенсорными и обеспечивают поступление в спинной мозг информации об активации рецепторов разных типов (табл. 3).
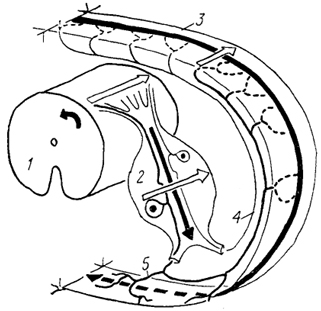
Рис. 16. Схема организации спинномозгового ганглия кошки (Kausz, Rethelyi, 1985).
1 - спинной мозг; 2 - ганглий; 3 - стенка туловища; 4, 5 - дорсальная (4) и вентральная (5) ветви чувствительного нерва. Стрелки показывают соответствие зон иннервации положению клеток в ганглии и спинном мозге.
Расположение клеток, связанных с разными видами чувствительности и иннервирующих определенные участки тела, строго упорядочено. Так, у кошки дорсовентральный градиент распределения зон иннервации в коже соответствует проксимальнодистальному градиенту распределения клеток в ганглии, а каудальноростральный градиент зон иннервации - медиолатеральному градиенту распределения клеток (рис. 16).
На сегментарном уровне организации спинного мозга сохраняется закономерный характер проекций. Во-первых, на уровне каждого сегмента по афферентным волокнам всех типов в спинной мозг поступает информация от строго определенного участка тела. Во-вторых, волокна, иннервирующие более дистально расположенные участки тела имеют намного меньшие рецептивные поля, чем проксимальные. Этим обеспечивается более высокая точность различения сигналов от дистальных участков конечностей (пальцы), связанных, как правило, с выполнением более тонких и дифференцированных движений, чем от проксимальных отделов конечностей.
Развитие афферентных связей спинного мозга в филогенезе позвоночных
Наиболее подробно у позвоночных изучено взаимодействие первичных афферентных волокон с мотонейронами спинного мозга. У круглоротых, как указывалось выше, часть клеток Рогон-Берда и клеток спинальных ганглиев не только устанавливает контакты с интернейронами, но и активирует мотонейроны посредством образования аксо-соматичеcких синапсов электро-химического типа.
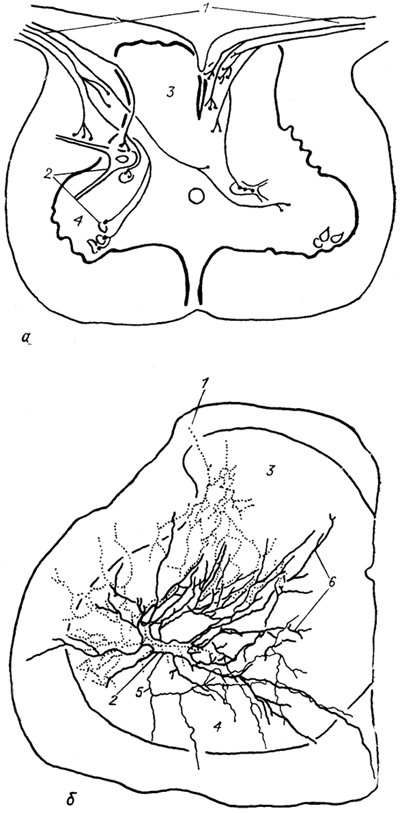
Рис. 17. Окончание афферентных волокон в спинном мозге карпа (а) и на" одиночном мотонейроне спинного мозга лягушки (б) (Моторина, 1982; 1983).
1 - афферентные волокна и их ветви; 2 - мотонейроны; 3,4 - задние (3) и передние (4) рога серого вещества; 5, 6 - аксон (5) и дендрит (6) мотонейрона.
У рыб выявлены афферентные волокна двух типов. Одни из них оканчиваются на интернейронах медиальной части дорсального рога серого вещества спинного мозга, другие образуют аксо-соматические и аксо-дендритные контакты смешанного типа на мотонейронах вентрального рога. Часть афферентных волокон может переходить на противоположную сторону спинного мозга (рис. 17). У ряда изученных хрящевых рыб (например, некоторые скаты из группы Rajaformes и акулы Squalus) обнаружены прямые моносинаптические связи с мотонейронами, тогда как у других (например, акулы Scyliorhinus canicula) таких связей не выявлено.
У амфибий афферентные волокна образуют контакты не только с дистальными дендритами мотонейронов (как у большинства рыб), но и с их проксимальными ветвями и телами. Число контактов отдельного волокна с мотонейроном может достигать 60 - 70 (рис. 17). Показано, что афферентные волокна от определенной мышцы ветвятся среди большой группы мотонейронов, связанных с иннервацией различных мышц, но образуют контакты только с мотонейронами "своей" мышцы. У амфибий в спинном мозге, впервые среди позвоночных, намечается четкое топографическое разделение зон ветвления афферентов разного типа. Первичные афференты мышечных волокон (тип А) контактируют с мотонейронамн в вентральном. роге, тогда как афференты кожной и мышечной чувствительности (типы Iа, II) связаны с мотонейронами через систему интернейронов, и зона их ветвления ограничена дорсальным рогом.
Распределение афферентов в спинном мозге рептилий сходно с таковым у амфибий. Часть волокон оканчивается в дорсальном роге, другие достигают вентральных рогов. Электро-физиологически показано наличие прямых моносинаптических связей афферентов с мотонейронами у ящериц, крокодилов и черепах.
У млекопитающих и птиц происходит дальнейшая дифференциация входов. Основная масса проекций от кожных рецепторов (связанных с ноци-, механо- и терморецепцией) направляется в ядра дорсального рога серого вещества, а проекции таких специализированных рецепторов, как сухожильные, расположены в основании дорсального рога. Таким образом, большинство афферентов не достигает вентральных рогов спинного мозга, их контакт с мотонейронами осуществляется полисинаптически, через систему интернейронов (рис. 18). Ветвления отдельных афферентных волокон строго упорядочены и формируют сложную систему пространственных модулей, обеспечивающих высокий уровень специфичности межнейронных связей.
Серое вещество спинного мозга состоит из тел и отростков нейронов и у всех позвоночных расположено вокруг центрального спинномозгового канала, являющегося остатком полости
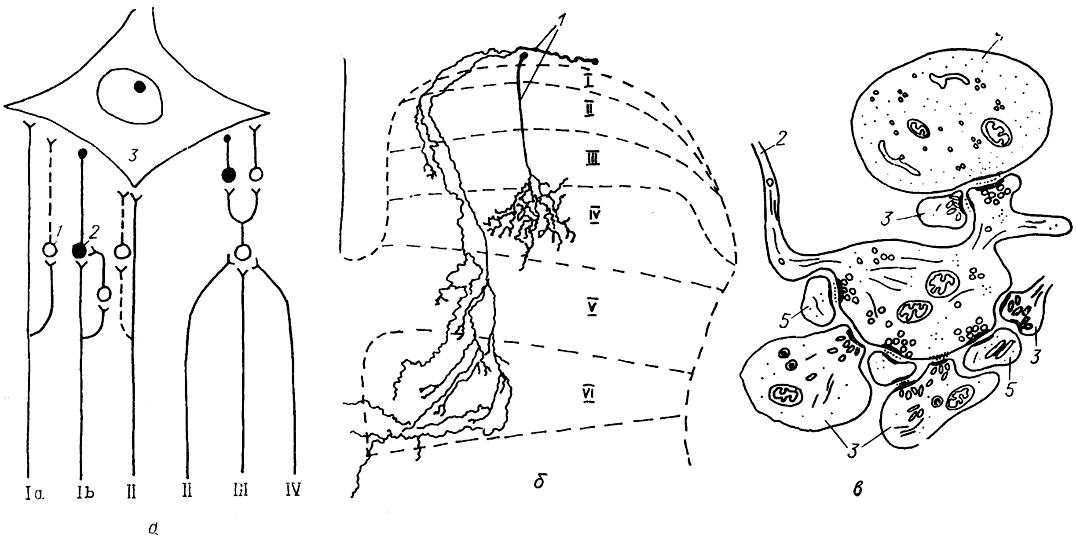
Рис. 18. Схема окончания первичных афферентов на мотонейроне (а), характер их ветвления в сером веществе задних рогов (б) и фрагмент ультраструктуры афферентного окончания (в) (Mannen, 1978; Honk, Zevrymer, 1981; Semba, 1983).
a: Ia, Ib, II-IV-различные виды афферентов; 1, 2 - возбуждающие (1) и тормозные (2) интернейроны; 3 - мотонейрон. б, в:1 - афференты; 2, 3 - аксонные терминали афферентов типа Аα (2) и Аβ (3); 4, 5 - дендрит (4) и дендритный шипик (5) интернейронов задних рогов; I-VI - пластины Рекседа.
нервной трубки. Основная эволюционная тенденция в развитии серого вещества состоит, во-первых, в миграции клеток от центрального канала на периферию и, во-вторых, в прогрессивной дифференциации клеточных популяций на отдельные зоны и ядра. Так, у бесчерепных все клетки спинного мозга располагаются вокруг центрального канала, тогда как у круглоротых часть нейронов активно мигрирует в толщу спинного мозга, формируя две латерально расположенные группы. У остальных позвоночных в спинном мозге образуются отдельные популяции нейронов дорсальных и вентральных рогов. Позднее, у наземных позвоночных, в ряде отделов спинного мозга появляются боковые рога.
В 1952-1954 гг. Б. Рексед предложил новую универсальную классификацию серого вещества спинного мозга, разделив его на десять пластин (I-X), различающихся по структуре составляющих их элементов и по связям (рис. 15, а).
Пластина I представлена узкой полоской нейронов с характерными тангенциально ориентированными дендритами. На интернейронах пластины I оканчиваются афференты типов Аβ, Аδ и С. Аксоны интернейронов образуют восходящие тракты спинного мозга, а также посылают коллатерали в более глубокие слои спинного мозга (рис. 19). Помимо сенсорных волокон в I пластине обнаружены многочисленные афференты от вышележащих отделов спинного и головного мозга, в том числе от структур лимбической системы, через которые осуществляется контроль болевых ощущений.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|