- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 4 страница
Пластины II и III построены из нейронов среднего размера, дендриты которых могут проникать как в зону пластины I, так и в более глубокие слои серого вещества. Аксоны этих клеток распространяются на большие расстояния и захватывают несколько сегментов, образуя многочисленные внутри- и межсегментные связи. Часть аксонов образует систему восходящих трактов.
Обилие входящих в пластины I-III афферентных волокон, разнообразие нейронных типов, наличие синоптических комплексов конвергентного и дивергентного типов и модуляторных контактов определяют сложный характер межнейронных взаимодействий. Так, в вентральной части пластины III особенно многочисленны специфические формы аксо-аксональных синапсов между нейронами и афферентами, а также синапсы дендро-дендритного типа (рис. 18). Таким образом, значение нейронных комплексов пластин I-III состоит в контроле поступающей в спинной мозг сенсорной информации различного вида, в ее переработке и передаче в спинной и головной мозг.
Пластины IV-VI представлены скоплениями нейронов мультиполярного типа, дендриты которых ориентированы преимущественно в дорсальном направлении и проникают в зону пластин I-III. На телах и дендритах нейронов оканчиваются
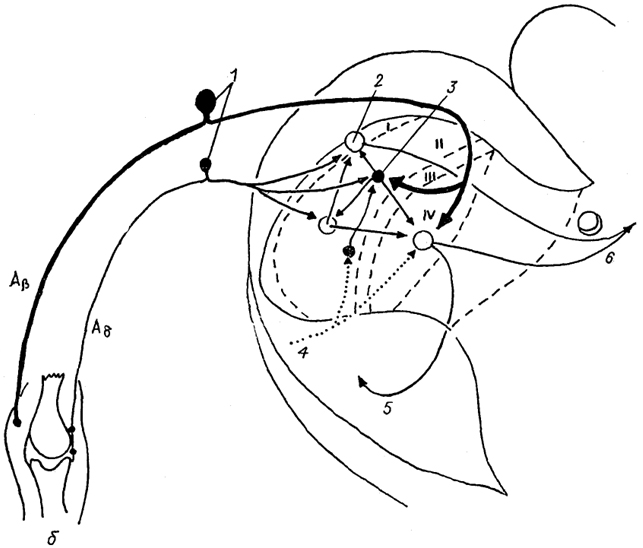
Рис. 19. Межнейронные связи в I-IV пластинах заднего рога спинного мозга (Delwaide, Schoenen, 1989).
Aβ, Aδ - афференты; 1 - чувствительные нейроны спинального ганглия; 2, 3 - возбуждающие (2) и тормозные (3) интернейроны; 4 - нисходящий бульбо-спинальный тракт; 5, 6 - восходящие спинно-ретикуло-таламический (5) и спинно-таламический (6) тракты.
многочисленные терминали первичных афферентов из групп Аα, Аβ, Аδ, аксоны интернейронов собственного и соседних сегментов, а также волокна нисходящих трактов. Окончания афферентов строго упорядочены: в вентральный отдел пластины V проецируются болевые и высокопороговые механорецепторы, в средний - рецепторы кожи, в дорсальный - низкопороговые: механорецепторы волосяных фолликулов. Эти отделы спинного мозга являются источником основной массы проприоспинальных (собственных) путей спинного мозга, образуя три системы связей: короткие - в пределах 1-2 сегментов, промежуточные- 4-12 сегментов, и Длинные. Последние представлены аксонами, идущими в составе тонкого пучка Голля в продолговатый мозг, в то время как их коллатерали достигают промежуточного мозга. Нейроны, аксоны которых образуют систему длинных связей, локализованы в основном в пластине IV шейных и пластинах V и VI пояснично-крестцовых сегментов спинного
мозга. Промежуточные и короткие проприоспинальные, пути связаны с проведением информации от рецепторов мышц, суставов, сухожилий. Часть волокон оканчивается на крупных нейронах ядра Кларка в пластине V, образующих в свою очередь крупный восходящий дорсальный спинно-мозжечковый тракт. Нейроны ядра Кларка получают проекции от афферентов флексорного рефлекса (FRA), связанных с иннервацией конечностей, причем разные афференты проецируются на различные клетки этого ядра.
Пластины VII и VIII, как полагают, не получают прямых афферентных проекций и занимают промежуточное положение между задними и передними рогами серого вещества спинного мозга. Здесь широко представлены терминальные ветвления проприо- и супраспинальных трактов. Аксоны интернейронов VII и VIII пластин оканчиваются на мотонейронах и образуют восходящие и нисходящие тракты различного типа. Одной из разновидностей этих интернейронов являются клетки Реншоу, получающие коллатерали аксонов мотонейронов и осуществляющие обратное торможение последних. Латеральные отделы пластины VII на уровне грудных - верхних поясничных сегментов образуют боковые рога и состоят из скоплений ассоциативных преганглионарных нейронов вегетативной нервной системы. Современные исследования, проведенные с помощью внутриклеточных инъекций пероксидазы хрена, показали сложную структурно-функциональную организацию боковых рогов. Их нейроны относятся к ретикулярному типу, для которого характерно широкое распространение дендритав, достигающих даже пластины I задних рогов. На дендритах и телах преганглионарных нейронов обнаружены многочисленные контакты различного происхождения. В целом подробный анализ нейронной структуры промежуточной зоны спинного мозга показал, что это образование ретикулярного типа, переходящее непосредственно в ретикулярную формацию ствола головного мозга и являющееся связующим звеном между сенсорными формациями заднего рога и моторными ядрами переднего рога.
Вентральные рога спинного мозга представляют особый интерес, так как здесь располагаются группы спинальных мотонейронов, управляющих деятельностью поперечнополосатой скелетной мускулатуры. В спинном мозге млекопитающих мотонейроны занимают район пластин VIII и IX и дифференцированы на группы ядер: медиальную, центральную и латеральную (рис. 15, а). Мотонейроны являются наиболее крупными клетками спинного мозга и разделяются на три группы: α-мотонейроны, крупные и обеспечивающие быстрые сокращения мышц; β-мотонейроны, мелкие и обеспечивающие медленные сокращения мышц, и γ-мотонейроны, связанные с иннервацией мышечных веретен. На примере организации моторных ядер наиболее отчетливо проявляется принцип соматотопии: каждая
мышца или группа мышц иннервируется определенной группой мотонейронов (мотонейронный пул). В пределах сегмента организация мотонейронных пулов млекопитающих подчиняется так называемому правилу Романеса: нейроны, иннервирующие дистальные мышечные группы конечностей, располагаются в спинном мозге дорсально по отношению к нейронам, иннервирующим проксимальные мышечные группы. При этом мотонейроны мышц-сгибателей и -разгибателей, часто относящихся к одной мышечной группе, в структуре моторных ядер также располагаются изолированно друг от друга.
Пластина X - центральное серое вещество спинного мозга занимает небольшой участок вокруг центрального канала. Здесь располагаются мелкие интернейроны, связывающие ее с другими участками серого вещества, а также проходят пучки комиссуральных волокон, соединяющих правую и левую половины спинного мозга.
Эволюция мотонейронных центров спинного мозга позвоночных
У круглоротых в спинном мозге намечается дифференциация мотонейронов на популяции, иннервирующие отдельные миомеры. Каждая такая популяция включает в себя группы первичных (крупных) и вторичных (мелких) мотонейронов, иннервирующих внутри отдельного миомера различные зоны (дорсальную или вентральную) и типы мышц. Первичные и вторичные мотонейроны образуют множественные контакты (от 3 до 8) на волокнах быстрых "белых" мышц - полинейронная иннервация. Мононейронная иннервация, осуществляемая только первичными мотонейронами, характерна для медленных "красных" мышц. Однако у круглоротых нет обособления отдельных популяций мотонейроров, иннервирующих разные типы мышц в пределах сегмента.
У хрящевых рыб дифференциация белого и серого вещества в спинном мозге нечеткая (рис. 20). В вентральных рогах имеются три группы клеток: дорсомедиальная, вентролатеральная и центральная. Для мотонейронов характерна большая вариабельность формы и размеров, а также тенденция к тесному расположению их тел и отростков. Особенность мотонейронов рыб состоит в том, что они увеличиваются в размерах в течение всей жизни организма (в отличие от нейронов других позвоночных).
У акул и скатов мотонейроны, иннервирующие красные и "белые мышцы миомеров, уже пространственно разграничены (рис. 20, в, г). Красные мышцы иннервируются мелкими мотонейронами, сосредоточенными в латеральной части мотонейронного пула, белые - крупными мотонейронами, занимающими медиальные районы. Более того, аксоны мотонейронов, иннервирующие
разные участки миомеров, имеют разный диаметр и проходят в разных участках вентральных корешков.
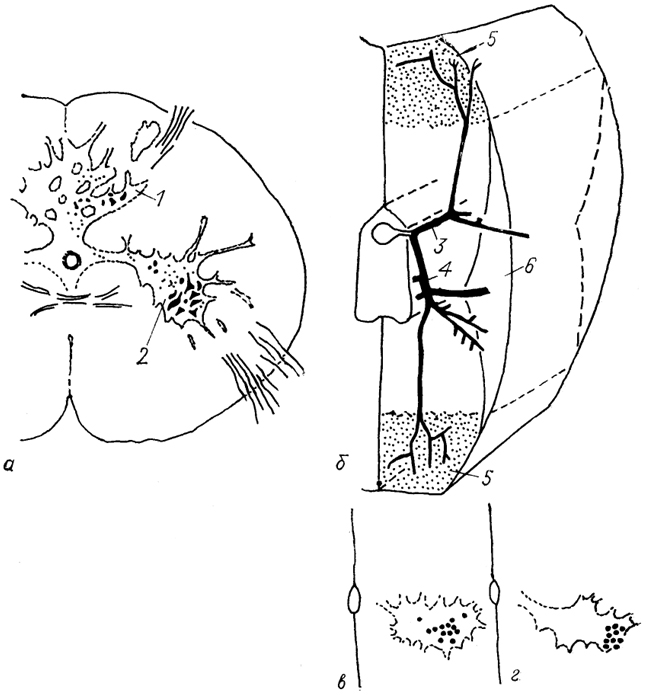
Рис. 20. Цитоархитектоника спинного мозга (а) , ветвление вентрального корешка спинномозгового нерва (б) и локализация мотонейронов, иннервирующих "красные" (в) и "белые" (г) мышцы у акулы (Mos, Willamson, 1986).
1, 2 - задние (1) и передние (2) рога серого вещества; 3, 4 - дорсальная (3) и вентральная (4) ветви вентрального корешка спинномозгового нерва; 5, 6 - зона "белых" (5) и "красных" (6) мышц миотома.
Мотонейроны костистых рыб расположены в вентральных рогах серого вещества спинного мозга и представлены двумя популяциями. Первую популяцию составляют крупные первичные мотонейроны, локализованные исключительно в дорсальной части мотонейронной колонны. Их сильно развитые дендриты проникают в белое вещество и часто достигают противоположной стороны спинного мозга. Таких мотонейронов немного: например, у золотой рыбки на 1/2 сегмента приходится 8-12
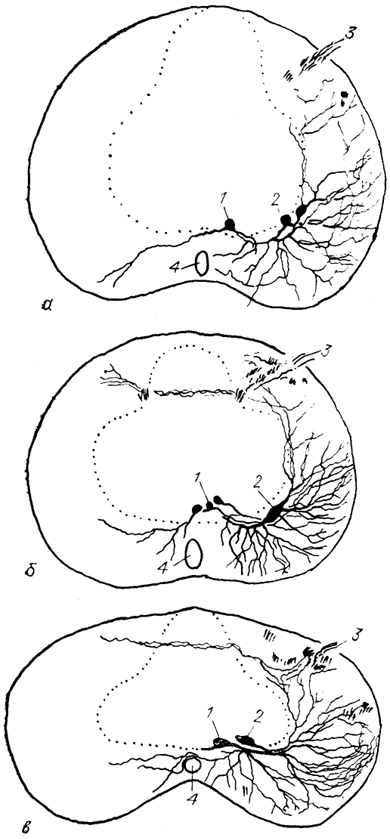
Рис. 21. Нейронная структура моторных ядер шейных сегментов спинного мозга хвостатых амфибий из группы саламандр: Rhyacotriton olympicus (a), Ptethodon jordani (б), Batrachoseps attenuatus (в) (Toth e. a., 1985).
1, 2 - медиальная (1) и латеральная (2) группы мотонейронов; 3 - дорсальный корешок; 4 - спинномозговой канал. Точками обозначена зона серого вещества.
клеток. Они образуют аксо-аксональные контакты электротонического или смешанного типа с маутнеровскими нейронами. Вторая популяция представлена мелкими вторичными мотонейронами, занимающими в мотонейронной колонне сегмента вентральное положение. Отдельные первичные мотонейроны иннервируют определенные участки миомеров, а разные популяции мотонейронов иннервируют в миомере различные типы мышц: первичные - белые, а вторичные - и белые и красные мышцы. Отдельное мышечное волокно иннервируется несколькими мотонейронами - полинейронная иннервация. Электрические органы, имеющиеся у некоторых рыб (в частности, у электрических скатов, сомиков) и развивающиеся из производных осевой мускулатуры, иннервируются специальной популяцией мотонейронов, расположенных в центральной зоне спинного мозга и связанных друг с другом многочисленными электротоническими контактами сомато - соматического и дендро-дендритного типов.
У амфибий, как и
у других низших позвоночных, мотонейроны разделяются на два основных класса: первичные (крупные) мотонейроны, дендриты которых распространяются на большие расстояния, захватывая несколько сегментов и переходя на противоположную сторону спинного мозга, и вторичные (мелкие) мотонейроны, аксоны которых не контактируют с маутнеровскими волокнами.
У хвостатых амфибий популяции мотонейронов формируют две группы, или колонны: вентромедиальную и вентролатеральную. Первая содержит в основном более мелкие грушевидные клетки, а вторая представлена крупными веретеновидными нейронами, причем степень развития той или иной мотонейронной колонны сильно варьирует даже у представителей одной труппы животных (рис. 21). Иннервация мышц у хвостатых амфибий полинейронная, и популяции мотонейронов содержат как первичные, так и вторичные мотонейроны.
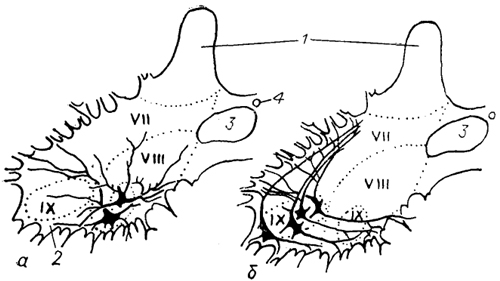
Рис. 22. Нейронная структура вентромедиального (а) и вентролатерального (б) мотонейронного ядра спинного мозга ящерицы (Cruce, 1979).
1, 2 -задние (1) и передние (2) рога спинного мозга; 3 - медиальные проприоспинальные тракты; 4 - спинномозговой канал; VII-IX -пластины Рекседа.
У бесхвостых амфибий в спинном мозге намечается пространственное разделение мотонейронных популяций, иннервирующих разные группы мышц. Клетки, иннервирующие осевую туловищную мускулатуру, расположены в мотонейронной колонне сегмента вентральнее и медиальнее мотонейронов, иннервирующих конечности, хотя внутри пула еще нет морфологической дифференцировки нейронов, иннервирующих отдельные мышцы. Мотонейроны образуют многочисленные электротонические, химические и смешанные контакты с клетками соседних сегментов и противоположной стороны спинного мозга, создавая основу для координированной деятельности мышц туловища и конечностей.
Мотонейроны спинного мозга рептилий организованы в две
моторные группы: вентромедкальную и вентролатеральную, нейроны которых отличаются друг от друга по размеру и характеру ветвления дендритов (рис. 22). Мотонейроны первой группы иннервируют осевую мускулатуру, второй - мускулатуру конечностей. У рептилий мотонейроны, иннервирующие отдельные мышцы, уже расположены раздельно. Однако в связи с тем, что у них самая сложная среди позвоночных мышечная система, имеется множество вариантов моторной иннервации. Так, красные медленные мышечные волокна рептилий иннервируются полинейронально и вдоль всей поверхности волокна, тогда как белые фазные волокна, разделяемые на быстрые и медленные, иннервируются мононейронально и локально. Очень часто в отдельных мышцах красные и белые волокна перемешаны друг с другом и иннервируются разными мотонейронами. Считается, что такая иннервация мышц у рептилий обеспечивает синхронизацию их работы (например, при движении туловища у змей). Подчеркнем, что у рептилий впервые среди позвоночных дендриты большинства мотонейронов не проникают на противоположную сторону спинного мозга, и это позволяет осуществлять более тонкую регуляцию деятельности мотонейронов спинномозгового сегмента через систему интернейронов.
Организация мотонейронных популяций, иннервирующих осевую мускулатуру млекопитающих, подробно изучена у крыс и кошек. Мотонейроны, связанные с определенными мышцами, занимают строго определенное положение в мотонейршном пуле. При этом мышцы, которые развиваются в соседних участках миомеров, иннервируются рядом расположенными нейронами, несмотря на возможное изменение положения данных мышц в туловище взрослого животного. У млекопитающих γ-мотонейроны, связанные с интрафузальными мышечными волокнами, рассеяны среди других нейронов данного пула. Интересно, что часть осевой мускулатуры млекопитающих - шейные мышцы - сохраняет филогенетически древний принцип иннервации - двумя разными типами мотонейронов, расположенных в разных ядрах. Остальная, как быстрая белая, так и медленная красная мускулатура не имеет двойной иннервации. Таким образом, в процессе эволюции сохраняются все возможные варианты организации нервно-мышечных отношений и соответственно мотонейронных пулов спинного мозга, причем часто независимо от филогенетического уровня.
§ 2. ПРОВОДЯЩИЕ ПУТИ СПИННОГО МОЗГА
Все проводящие пути спинного мозга сосредоточены в белом веществе, разделяемом на три канатика: задний., боковой и передний. Основной объем канатиков занимают супраспи-нальные тракты, осуществляющие двустороннюю связь спинного
и головного мозга. Тракты, занимающие узкую полоску вокруг серого вещества, называются проприоспинальными (рис. 15).
Проприоспинальные тракты образованы аксонами интернейронов промежуточной зоны серого вещества. Распределение тел и окончаний проприоспинальных нейронов у позвоночных, начиная с самых ранних этапов эволюции, строго упорядочено. У высших позвоночных аксоны нейронов латеральных отделов пластин V-VII проходят в боковом канатике и оканчиваются в дорсолатеральных областях латеральных моторных ядер, ин-нервирующих мышцы-сгибатели. Нейроны центра пластины VII проецируются в вентромедкальную область этих ядер, связанную с мышцами-разгибателями. Интернейроны из медиальных районов пластин VII и VIII проецируются в медиальные моторные ядра, которые иннервируют осевую туловищную мускулатуру. При этом нейроны, образующие длинные восходящие и нисходящие проприоспинальные волокна, располагаются преимущественно в вентральных участках пластин V-VIII, а их аксоны оканчиваются в вентромедиальных районах пластин V-VII.
Проприоспинальные нейроны, относящиеся к разновидности ретикулярных изодендритных нейронов, создают морфологическую основу для широкой конвергенции на их телах и дендритах супраспинальных и сенсорных афферентов. Большинства проприоспинальных нейронов активируется супраспинальными трактами моносинаптически, а первичными афферентами - полисинаптически. Таким образом, появление и развитие в эволюции позвоночных системы проприоспинальных межсегментарных связей обеспечивает (наряду с другими механизмами) координацию сложных моторных реакций.
Супраспинальные тракты наибольшего развития и сложности достигают у высших млекопитающих - приматов, у которых описано их более двух десятков (рис. 15). Остановимся на наиболее важных из них.
Дорсальные канатики содержат восходящие тракты, волокна которых образованы аксонами сенсорных нейронов и коллатералями интернейронов пластин V и VI. Система дорсальных канатиков у высших позвоночных построена по соматотопическому принципу: чувствительные волокна от нижней части туловища и нижних конечностей проходят медиально, формируя тонкий пучок Голля (fasciculus gracilis), от верхней части тела и верхних конечностей - латерально, образуя клиновидный пучок Бурдаха (fasciculus cuneatus), причем в самих трактах пучки волокон от определенных участков тела и конечностей проходят также отдельно друг от друга. Функционально они тоже разделяются: волокна, несущие информацию от рецепторов волосяных фолликулов и конечностей, составляют поверхностную часть пучков, от мышечных рецепторов – среднюю
часть, а от рецепторов давления и вибрации - глубокую центральную часть. Оканчиваются волокна дорсальных канатиков в соответствующих ядрах продолговатого мозга.
В эволюции позвоночных происходит морфологическое обособление дорсальных канатиков от латеральных. Так, уже у хрящевых и некоторых костистых рыб они разделяются волокнами нисходящего бульбо-спинального тракта. Волокна дорсальных канатиков достигают продолговатого мозга и оканчиваются в зоне, которая по своей структурно-функциональной организации соответствует ядрам этого тракта у высших позвоночных. У наземных позвоночных уровень дифференцировки дорсальных канатиков тесно связан с развитием конечностей. Например, среди рептилий он наиболее, высок у крокодилов и наименее - у змей. Кроме того, начиная с амфибий, в организации этой системы все более отчетливо проявляется соматотопический принцип, Достигающий своего максимального выражения у млекопитающих и человека.
Система восходящих путей латеральных канатиков включает в себя ряд крупных трактов, соединяющих спинной мозг с продолговатым - спинно-ретикулярные, со средним - спинно-тектальные, с промежуточным мозгом - спинно-таламические, и мозжечком - спинно-мозжечковые. У низших позвоночных еще нет анатомического разделения трактов. У рептилий латеральные канатики уже дифференцированы на дорсо- и вентролатеральные пучки. Первый содержит в основном спинно-ретикулярные, спинно-тектальные и спинно-мозжечковые тракты, а второй - спинно-таламические и большую часть спинно-ретикулярных трактов.
Спинно-ретикулярные тракты обнаружены у всех изученных позвоночных - от круглоротых до человека. Они оканчиваются в каудальных отделах ретикулярной формации продолговатого мозга, моторных ядрах черепномозговых нервов, достигая в ряде случаев ретикулярной формации среднего мозга. Считается, что спинно-ретикулярные тракты позвоночных обеспечивают проведение информации от болевых рецепторов, а также пролриорецепторов.
Спинно-тектальные тракты описаны у большинства позвоночных, за исключением миног, лучеперых рыб и бесхвостых амфибий (возможно, что они утратили их в процессе эволюции независимо друг от друга). Спинно-тектальные тракты оканчиваются в центральном сером веществе мозга, в глубоких слоях тектума, тегментуме и интерколликулярном ядре.
Спинно-таламические тракты (спинно-таламический и спинно-ретикуло-таламический) обеспечивают болевую и температурную чувствительность и проведение информации от волосяных фолликулов, рецепторов давления и мышечных афферентов III группы. Спинно-ретикуло-таламичесюий тракт, как филогенетически наиболее древний, описан практически у всех
позвоночных, тогда как прямой спинно-таламический тракт обнаружен лишь у амниот и хрящевых рыб. Объем спинно-таламического тракта, а также локализация нейронов, образующих его, место окончания и количество передаваемой информации значительно варьируют даже у близкородственных групп позвоночных. Так, у млекопитающих проекции в специфические зоны таламуса (вентробазальный комплекс ядер) образованы нейронами пластин I и V спинного мозга, а в неспецифические зоны (медиальный комплекс ядер) -нейронами VII и VIII пластин. У птиц количество спинно-таламических нейронов очень невелико, полностью отсутствуют проекции из I пластины, при этом практически отсутствуют спинно-таламические проекции из шейных сегментов спинного мозга, иннервирующих крылья.
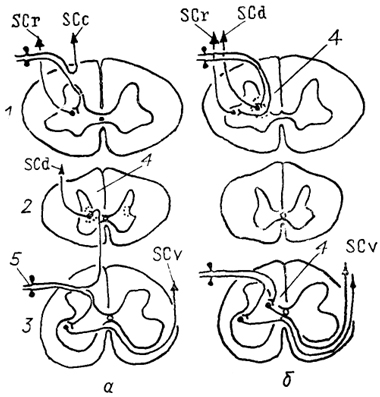
Рис. 23. Организация спинно-мозжечковых трактов у млекопитающих (а) и птиц (б) (Necker, 1989).
1-3 - шейный (1), грудной (2) и поясничный (3) отделы спинного мозга; 4 - ядро Кларка; 5 - чувствительные нейроны спинномозгового ганглия. Остальные обозначения, как на рис. 15.
Спинно-мозжечковые тракты есть у всех позвоночных, за исключением миног и миксин. У низших позвоночных спинно-мозжечковые волокна вместе с другими восходящими трактами входят в состав латерального канатика белого вещества спинного мозга и отделяются от них только в продолговатом мозге на уровне ядра V нерва. Разделение на вентральный и дорсальный компоненты намечается уже у хрящевых рыб и завершается у млекопитающих у которых выделяют дорсальный, вентральный и ростральный спинно-мозжечковые тракты и спинно-кунео-мозжечковый тракт (рис. 23). Дорсальный тракт начинается от крупных нейронов ядра Кларка в пластине IV серого вещества, оканчивается в I-IV и частично в V дольках мозжечка и несет информацию от отдельных мышц туловища и нижних конечностей. Вентральный образован нейронами нижних сегментов спинного мозга, расположенными в пластине V, и связан с проведением информации от группы мышц. Ростральный спинно-мозжечковый и спинно-кунео-мозжечковый тракты получают информацию от рецепторов верхних конечностей
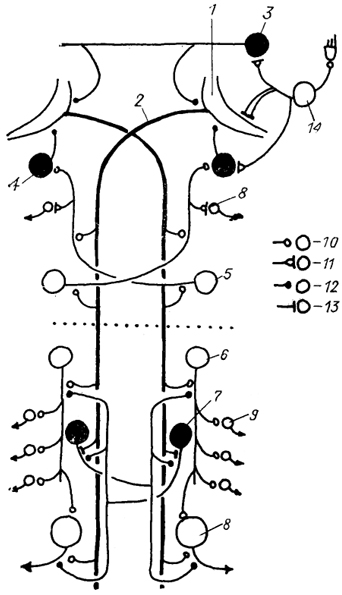
Рис. 24. Схема нейронной сети маутнеровских нейронов в спинном мозге костистых рыб (Faber e. а., 1989).
1 - маутнеровская клетка и ее аксон(2); 3-5 - комиссуральные (3) ,коллатеральные (4) и краниальные (5) интернейроны продолговатого мозга; 6, 7 - нисходящие (6) и реципрокные (7) интернейроны спинномозгового сегмента; 8, 9 - первичные (8) и вторичные (5) мотонейроны; 10-13 - синаптические контакты химического (10), электрохимического (11), химического тормозного (12) и электротонического (13) типов; 14 - нейроны чувствительного ганглия вестибулярного нерва. Темные клетки - тормозные, светлые - возбуждающие. Пунктиром обозначена граница головного и спинного мозга.
и начинаются в шейных сегментах спинного мозга. Все тракты и особенно дорсальный имеют четкую соматотопическую организацию. У птиц организация спинно-мозжечковых трактов иная, чем у рептилий и млекопитающих (рис. 23, б).
Нисходящие супраспинальные тракты опосредуют влияние головного мозга на моторные центры спинного мозга, и их активность связана прежде всего с такими рефлекторными актами, как плавание, бег, шагание, прыжки, полет. Система нисходящих трактов сформировалась на ранних этапах эволюции позвоночных и имеет единый план строения, включая несколько основных трактов: ретикуло-, вестибуло-, текто- и рубро-спинальные.
У низших позвоночных основную массу нисходящих трактов составляют ретикуло-спинальные тракты. Среди нейронов, образующих эти тракты, особо выделяются мюллеровские и маутнеровские клетки, расположенные в стволе мозга и обладающие длинными и нисходящими аксонами, которые моносинаптически связаны с мотонейронами спинного мозга, управляющими движением туловища и хвоста. У костистых рыб аксон каждой маутнеровской клетки ипсилатерально образует химические возбуждающие синапсы на спинальных нейронах двух типов: первичных мотонейронах и так называемых нисходящих интернейронах, аксоны которых на протяжении двух и более сегментов контактируют с первичными и вторичными мотонейронами (рис. 24).
Анализ структуры ретикуло-спинальных трактов высших позвоночных
показал, что данная система является наиболее консервативной. Это подтверждается ранним формированием ретикуло-спинальных трактов в онтогенезе.
Вестибуло-спинальные тракты развиты практически у всех позвоночных и разделяются на несколько пучков. У млекопитающих описаны прямой тракт (из латерального вестибулярного ядра) и два перекрещенных (из латерального и нисходящего вестибулярных ядер). Стимуляция этих ядер оказывает билатеральное действие на нейроны спинного мозга, связанные с движением головы, плечевого пояса и верхних конечностей. Тракт заканчивается на мотонейронах или интернейронах пластин VI-VIII.
Текто-спинальные тракты берут начало из различных отделов tectum opticum и опосредуют его влияние на мотонейроны спинного мозга. У млекопитающих они частично перекрещены. Обширные текто-спинальные тракты описаны у хрящевых и двоякодышащих рыб, амфибий (хвостатых и бесхвостых) и рептилий. По-видимому, они отсутствуют у некоторых костистых рыб (например, у лососевых). У круглоротых (по крайней мере, у миног) текто-спинальные волокна достигают верхних сегментов спинного мозга.
Перекрещенный рубро-спинальный тракт, сформированный нейронами красного ядра, обнаружен у бесхвостых и хвостатых амфибий, рептилий, птиц и млекопитающих; он, вероятно, отсутствует у безногих амфибий, змей и рыб. Прямой рубро-спинальный тракт описан у некоторых хрящевых (например, у скатов) и костных рыб, которые используют свои парные грудные придатки для передвижения. Таким образом, рубро-спинальные тракты эволюционно связаны с появлением конечностей и поясов конечностей, и степень их развития коррелирует со степенью развития мозжечка (однако присутствие рубро-спинального тракта у змеи Nerodia fasciata опровергает мнение о том, что он может быть связан только с регуляцией движений конечностей). Заканчиваются тракты в спинном мозге на интернейронах пластин V-VII.
Кортико-спинальный (пирамидный) тракт появляется у птиц и млекопитающих и является наиболее молодой из всех проводящих систем спинного мозга. Размер тракта увеличивается в ряду млекопитающих, тесно коррелируя с развитием моторной активности вида и со способностью к тонким дифференцированным движениям конечностей, и достигает максимума у человека. В спинном мозге зона прямых проекций тракта значительно варьирует у разных видов (рис. 25). У низших млекопитающих (например, у сумчатых) кортико-спинальные волокна достигают только шейных и грудных сегментов, проходя в дорсальных канатиках, и оканчиваются в задних рогах спинного мозга. У высших млекопитающих кортико-спинальный
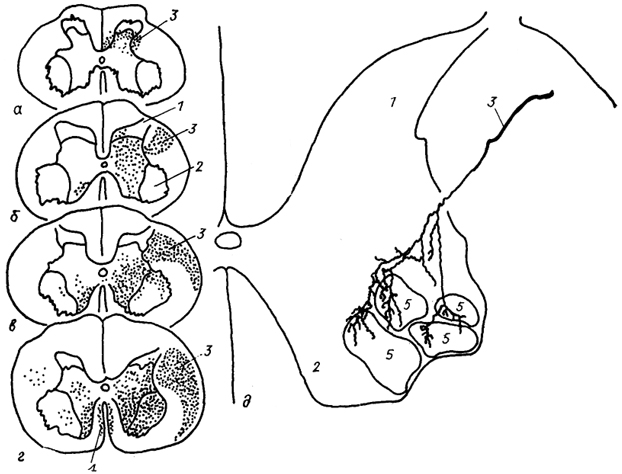
Рис. 25. Окончание кортико-спинального тракта в шейных сегментах спинного мозга опоссума (а), кошки (б), макаки (в), шимпанзе (г) и характер ветвления одного волокна в моторных ядрах мозга макаки (д) (Shinoda е. а., 1979; Kuypers, 1981).
1, 2 - задние (1) и передние (2) рога спинного мозга; 3, 4 - латеральный (3) и передний (4) кортико-апинальные тракты; 5 - моторные ядра.
тракт проходит вдоль всего спинного мозга, разделяясь на два! компонента (латеральный и вентральный).
Кортико-спинальные волокна начинаются в разных областях новой коры, и, таким образом, пирамидный тракт по существу представляет собой комплекс отдельных путей, каждый из которых отличается по характеру окончаний в спинном мозге. Так, у высших приматов волокна от моторных зон коры проецируются в промежуточную зону и моторные ядра, волокна от соматосенсорных зон - в пластины I-IV. Дифференцированное распределение кортико-спинальных проекций свидетельствует об их влиянии на разные спинальные механизмы, вплоть до регуляции сенсорных афферентов в пластинах I и II. Более того, волокна из моторных зон коры оканчиваются преимущественно на телах и проксимальных дендритах мотонейронов, тогда как проекции из соматосенсорных полей - на дистальных дендритах. У большинства млекопитающих пирамидный тракт созревает в постнатальный период, причем в процессе его развития происходит элиминация многих кортико-спинальных
волокон, выходящих из некоторых корковых зон (например, зрительной коры).
Таким образом, уже в строении спинного мозга различных позвоночных отчетливо прослеживаются тенденции последовательной дифференциации его элементов и связей, а также усложнение межнейронных взаимодействий, обусловленные системным повышением уровня организации ЦНС.
ГЛАВА 3
ПРОДОЛГОВАТЫЙ МОЗГ
Продолговатый мозг, Medulla oblongata, уже у круглоротых представлен в достаточно дифференцированном виде. Являясь продолжением спинного мозга, он на значительном протяжении сохраняет свойственные последнему структурные особенности и форму. Изменение конфигурации и взаимного расположения структур продолговатого мозга у всех позвоночных происходит в дорсальных отделах в результате формирования IV желудочка мозга путем расщепления крыловидной пластинки и расхождения ее в вентролатеральных направлениях. Рострально желудочек переходит в полость среднего мозга. Верхняя его часть сформирована хориоидной крышей и у большинства позвоночных на всем протяжении ограничена мозжечком.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|