- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 10 страница
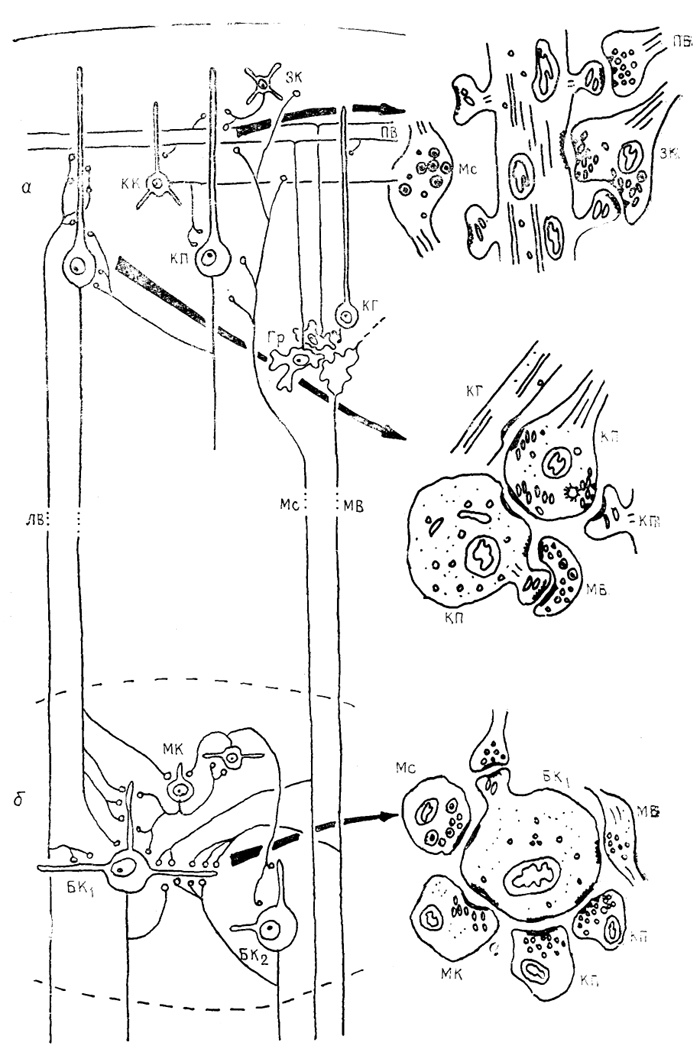
Рис. 47. Организация связей в мозжечке приматов (по: Chan-Palay, 1977)
а - кора, б - ядра мозжечка. МК - малые, БК1 и БК2 - большие клетки ядер мозжечка. Остальные обозначения, как на рис. 45, 46.
дают начало параллельным волокнам, идущим перпендикулярно плоскости ветвлений отростков звездчатых нейронов и клеток Пуркинье.
Клетки Гольджи - крупные нейроны гранулярного слоя, размеры их тел составляют у млекопитающих 11-17 мкм, у рептилий - 18-25 мкм. Дендриты некоторых из них ветвятся в пределах этого слоя, у других доходят до молекулярного. В отличие от большинства клеток мозжечка, отростки которых ветвятся в одной плоскости, дендриты клеток Гольджи формируют (по крайней мере, у млекопитающих) в микропространстве мозжечка нечто, похожее на цилиндр. Их аксоны также охватывают некий цилиндрический объем в пределах гранулярного слоя и устанавливают тормозные контакты с дендритами клеток-зерен.
Мшистые волокна, попадая в гранулярный слой, формируют по ходу розетки, от них начинаются короткие отростки, интер-дигитирующие с дендритами клеток-зерен. Этот комплекс образует мозжечковые гломерулы, которые на препаратах выглядят как области, лишенные клеточных тел. Наиболее сложно организованы гломерулы у высших млекопитающих, наиболее просто - у рептилий. К периферии каждой гломерулы подходят аксоны клеток Гольджи, которые в ответ на активацию параллельными или мшистыми волокнами оказывают тормозное действие. В центре гломерул располагаются терминали коллатералей лиановидных волокон. Гломерулы - характерные компоненты гранулярного слоя высших позвоночных - формируются при участии аксонов клеток Гольджи, коллатералей лиановидных волокон и розеток мшистых волокон. У млекопитающих каждое мшистое волокно образует до 16 розеток, каждая из которых формирует связи с 28 клетками-зернами. Таким образом одно мшистое волокно воздействует одновременно на 448 разных клеток-зерен. Коллатерали лиановидных волокон помимо гломерул образуют и контакты "по ходу" с телами клеток Гольджи.
Ганглионарный слой представлен наиболее крупными нейронами мозжечка - клетками Пуркинье, размеры их грушевидных тел составляют 20-25 мкм у млекопитающих, 25-35 мкм у рептилий. Гладкий начальный сегмент единственного дендрита дает начало гладким стволам вторичных и третичных дендритов. Тончайшие дендриты 3-го порядка особенно многочисленны у млекопитающих (у кошки - до 80 тыс.), и от них в свою очередь отходит множество ветвей, богатых шипиками. Арборизация дендритов клеток Пуркинье у рептилий ниже, чем у птиц и млекопитающих. Совокупность ветвлений практически располагается в одной, сагиттальной, плоскости, перпендикулярно оси долек, и, пронизывая всю толщу молекулярного слоя, достигает его поверхности. Аксоны клеток Пуркинье начинаются от базального полюса и адресованы ядрам мозжечка,
а от клеток Пуркинье, расположенных в дольке X,- ядрам вестибулярного комплекса. На пути их через гранулярный слой образуется небольшое число (больше у млекопитающих, меньше у птиц и рептилий) возвратных коллатералей,* адресованных либо соме и дендритам клеток Гольджи (при отжждении коллатерали в глубине гранулярного слоя), либо соме и проксимальным отделам дендритов самой клетки Пуркинье, или корзинчатым клеткам (если коллатераль отходит в поверхностных отделах).
Лиановидные волокна у всех высших позвоночных образуют характерные контакты с дендритами клеток Пуркинье: волокно, сильно ветвясь, сопровождает дендрит на всем его протяжении и заканчивается синапсами на шипиках. Для млекопитающих и птиц доказано происхождение лиановидных волокон из нижней оливы, для рептилий подобное заключение сделано, исходя из особенностей данных контактов. Показано, что одно лиановидное волокно может контактировать с несколькими (у млекопитающих до 10) клетками Пуркинье. В свою очередь каждая клетка Пуркинье получает вход лишь от одного волокна, которое устанавливает до 200-300 синаптических контактов.
Параллельные волокна пронизывают дендритные ветвления клеток Пуркинье и формируют множественные контакты с их шипиками (до 80 тыс. у кошек), а поскольку каждое волокно образует не более двух контактов на одной клетке, то соответственно одна клетка Пуркинье получает входы от нескольких десятков тысяч волокон. В свою очередь одно параллельное волокно адресуется многим клеткам Пуркинье. Таким образом, данное звено представляет собой систему с высоким уровнем как конвергенции, так и дивергенции. Отметим, что далеко не каждое параллельное волокно контактирует с клеткой Пуркинье. Подсчеты, проведенные на мозжечке кошки, показали, что контакты устанавливает лишь каждое пятое волокно из проходящих через данную область ветвлений.
Помимо возбуждающих клетки Пуркинье получают и тормозные входы. Они сформированы аксонами звездчатых и корзинчатых клеток, а также волокнами катехоламинергической природы, происходящими из голубого пятна. Звездчатые и корзинчатые клетки располагаются в молекулярном слое и их количество прогрессивно уменьшается по направлению к поверхности. Отростки этих клеток в основном распределены в границах слоя.
Звездчатые клетки сосредоточены главным образом в наружных двух третях молекулярного слоя. Они представлены мультиполярами с округлым телом диаметром 7-13 мкм у млекопитающих и 10-15 мкм у рептилий. Их отростки ветвятся
в той же плоскости, что и у клеток Пуркинье. Среди звездчатых клеток выделяют коротко- и длинноаксонные; первые располагаются ближе к поверхности. У клеток, лежащих в разных отделах, меняется ориентация ветвлений: у расположенных поверхностно отростки направлены в глубь слоя, у лежащих в средних отделах - в обе стороны, а у находящихся в глубине - к поверхности.
Корзинчатые клетки располагаются непосредственно над клетками Пуркинье и параллельно им. Диаметр их тела составляет 10-15 мкм. Дендриты отходят радиально в плоскости, перпендикулярной длинной оси дольки. Аксон, начинаясь от тела или одного из дендритов, направляется сагиттально и проходит над многими клетками Пуркинье (над 18-20 у высших млекопитающих и над 6-8 у других позвоночных). Эти аксоны, формирующие так называемые поперечные волокна, дают несколько коллатералей: восходящие (в молекулярный слой), продольные (параллельно оси дольки на расстоянии 4-6 клеток Пуркинье) и нисходящие (в ганглионарный слой). Восходящие и продольные коллатерали образуют контакты с дендритами ганглиозных клеток, а нисходящие-перицеллюлярные корзинки, оплетающие сому, и кисточковидные окончания на начальном сегменте аксона клетки Пуркинье. Столь специализированная организация корзинчатых клеток представлена в мозге птиц и млекопитающих. У рептилий же она довольно примитивна: у многих видов (змеи, многие черепахи) ветвления корзинчатых клеток невелики, а число клеток, с которыми одна корзинчатая клетка устанавливает контакты, не превышает 2-3.
Афференты мозжечка высших позвоночных имеют несколько общих источников, среди которых в первую очередь выделяют сенсорные образования зрительной системы - ядро базального оптического тракта, вестибулярный комплекс и соматосенсорные отделы. Последние представлены системой ядер тройничного нерва и спинным мозгом. Наибольшее число спинальных источников обнаружено для мозжечка млекопитающих, у которых имеются дорсальный и вентральный спинно-церебеллярный тракты, trr. spinocerebellaris dorsalis et ventralis, ростральный и цервикальный пути, а также вход от латерального клиновидного ядра, tr. cuneocerebellaris. У птиц спинальные входы прослежены от цервикальной области, однако данные электрофизиологических исследований свидетельствуют о наличии упорядоченных проекций и от других соматических зон. У, рептилий если судить по локализации источников в спинном мозге, имеются дорсальный и вентральный спинно-мозжечковые тракты, гомологичные одноименным трактам млекопитающих. Другие входы у них не обнаружены, поскольку отсутствуют структуры, гомологичные их источникам (латеральное клиновидное ядро и ядро цервикального тракта).
Среди других структур, проецирующихся в мозжечок, выделяют
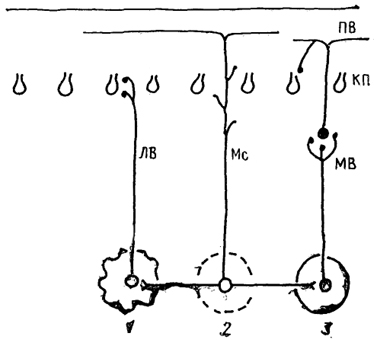
Рис. 48. Афференты коры мозжечка (по: Dietrichs, Haines, 1989).
1 - от нижней оливы, 2 - от гипоталамуса, 3 - от других отделов мозга. Остальные обозначения, как на рис. 45, 46.
группу так называемых прецеребеллярных ядер, основной объем эфферентов которых адресован мозжечку. К ним относятся: нижняя олива, ядра моста, тегментальное, парамедианное и латеральное (ядро латерального канатика) ретикулярные ядра. Что касается нижней оливы, то ее эфференты достигают мозжечка у всех высших позвоночных. Остальные в явном виде присутствуют в мозжечке птиц и млекопитающих. У рептилий (и, возможно, амфибий) существуют прекурсоры источников, локализованных в ретикулярной формации. Мостовые ядра у них не обнаружены, а у млекопитающих и птиц они составляют существенное звено в системе афферентов мозжечка. У млекопитающих ядра моста опосредуют передачу информации от коры большого мозга, а также, в меньшей степени, от тектума и спинного мозга. У птиц существуют гомологичные проекции от двух ядер, лежащих в пределах продолговатого мозга. На основании распределения этого вида проекций выделяют ту часть мозжечка, которая, будучи связанной с конечным мозгом, представляет собой наиболее молодой отдел-неоцеребеллум. У млекопитающих степень выраженности понтоцеребеллярных входов соответствует развитию моста как самостоятельного отдела мозга, и также уровню сложности мозжечка. Так, у полуобезьяны бушбеби, мозжечок которой имеет почти гладкую поверхность, и мост невелик. Сложный рельеф коры мозжечка у шимпанзе совпадает с крупным понтийным отделом. Мостовые проекции обеспечивают проведение импульсации преимущественно в контрлатеральные отделы коры мозжечка.
Еще одна группа афферентов характерна для всех высших позвоночных. Это серотонин- и катехоламинергические проекции от ядер шва и голубого пятна соответственно, адресованные коре мозжечка.
К специфическим особенностям афферентации мозжечка рептилий следует отнести многочисленные входы от ядра медиального продольного пучка и претектальной области. Для млекопитающих
и рептилий экспериментально подтвержден вход от гипоталамической области (рис. 48).*
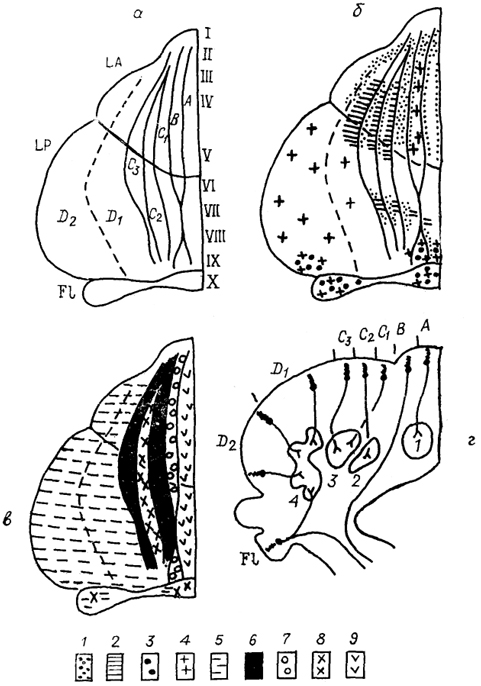
Рис. 49. Организация связей коры мозжечка приматов (Haines, 1986).
а - деление поверхности коры мозжечка на зоны A-D (объяснение в тексте) и на дольки 1-Х; LA - передняя, LP - задняя и F1 - флоккуло-нодулярная доли, б, в - распределение афферентов по коре мозжечка: 1 - спинно-мозжечковый, 2 - кунео-мозжечковый, 3 - вестибуло-мозжечковый тракты; 4 - первичные вестибулярные входы; 5-9 - проекции разных ядер нижнеоливарного комплекса: главной оливы (5), рострального (6) и каудального (7) полюсов дорсальной добавочной оливы, рострального (8) и каудального (9) полюсов медиальной добавочной оливы, г - эфференты коры мозжечка: показаны клетки Пуркинье и ход их аксонов к медиальному (1), заднему (2) и переднему (3) промежуточным и латеральному (4) ядрам мозжечка.
Существенной особенностью организации входов в мозжечок высших позвоночных является зональный характер распределения афферентов в коре. В наиболее оформленном виде это представлено у млекопитающих (рис. 49). Так, у приматов соматическая информация, поступающая по дорсальному спинно-мозжечковому тракту, закономерно распределяется по поверхности коры: туловищный отдел проецируется в медиальные участки долек II-V, конечности - в их латеральные участки, при этом задние конечности представлены в дольках II и III, передние - IV и V. Проекция области головы приходится на .дольку VI. Проекции, берущие начало от латерального клиновидного ядра, распределяются главным образом в области представительства передних конечностей (дольки IV и V) ив меньшей степени в каудальных .отделах. Входы рострального и вентрального спинно-мозжечковых трактов адресованы областям представительства передних и задних конечностей соответственно. Дополнительные соматосенсорные проекции обеспечиваются гюнто-мозжечковыми входами, которые опосредуют проведение импульсации от соматосенсорной области коры. В результате сложного, но закономерного распределения входов в коре мозжечка имеется не одна, описанная выше, а по крайней мере две области представительства. Вторая распределена в парамедиальной зоне VII-IX долек и организована так, что медиально располагается зона представительства туловища, латерально в рострокаудальном направлении - представительство головы, передних и задних конечностей. В мозжечке кошки также показана соматотопическая организация, причем ее характер достаточно сложен, в том числе из-за разной локализации зон проекций поверхностных и глубоких рецепторов.
Имеющиеся электрофизиологические данные позволяют заключить, что закономерным образом соматосенсорная информация представлена и в мозжечке птиц, у которых дольки I-IV связывают с представительством области хвоста и конечностей, дольки IV-VI - крыльев, а дольки V-VII - головы.
Закономерным образом распределены входы и от вестибулярного комплекса, причем данные, полученные на млекопитающих, показывают, что в отличие от ранее высказанных предположений область распределения этих окончаний не ограничивается дольками IX и X и срединной областью (область червя) - часть входов адресуется латеральным отделам коры. Входы от нижней оливы представлены лиановидными волокнами, которые распределяются в виде парасагиттально ориентированных полос. Ретикуло-мозжечковые проекции не имеют выраженной топики.
Для всех эфферентов коры мозжечка характерен зональный принцип распределения, который выражается в том, что определенным образом ориентированные клетки Пуркинье посылают свои аксоны в определенные участки мозжечковых ядер или
вестибулярного комплекса. У всех высших позвоночных выделяют три зоны: центральную (область червя, Vermis, у млекопитающих), паравермальную и латеральную. В случае четкого разделения X дольки выделяют и клочок, flocculus, как самостоятельную область. Дальнейшее изучение привело к уточнению зонального характера организации коры мозжечка. Так, у некоторых рептилий выделяют четыре зоны коры, у птиц - шесть, у млекопитающих - семь. Дополнительное разделение проводится в пределах обозначенных выше медиальной, латеральной, парамедиальной, или паравермальной зон (рис. 50).
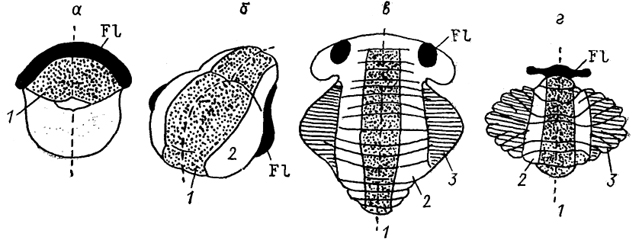
Рис. 50. Основные отделы мозжечка наземных позвоночных (Bangma, 1983).
а - амфибии, б - рептилии, в - птицы, г - млекопитающие. 1 - область червя, 2 - паравермальная и 3 - латеральная области.
У рептилий клетки Пуркинье, лежащие по средней линии, проецируются в медиальное ядро, клетки Пуркинье основной части промежуточной зоны - в вестибулярный комплекс (главным образом в дорсолатеральное и вентролатеральное ядра), небольшой участок промежуточной зоны (у черепах занимающий ростролатеральную, а у варанов - каудолатеральную часть) образует входы к латеральному ядру. Латеральная зона и флоккулус формируют проекции к нисходящему и вентромедиальному вестибулярным ядрам. У некоторых рептилий имеются отличия. Так, например, в мозжечке каймана не выделяется латеральная зона, а у питона от латеральной зоны имеются входы к латеральному ядру. У птиц паравермальная зона проецируется на группу клеток, гомологичную промежуточному ядру млекопитающих, латеральная зона - на латеральное ядро, а группа клеток, лежащая по средней линии, - на медиальное ядро.
У млекопитающих в состав медиальной зоны входят зона А и зона В, различающиеся по проекциям (рис. 49, г). Зона В в дольках VII и VIII сильно редуцирована или вовсе отсутствует. Зона Апроецируется на медиальное ядро, зона В - на вестибулярный комплекс. В составе промежуточной зоны в медиолатеральном направлении расположены зоны С1, С2, и С3,
проецирующиеся на медиальную часть переднего промежуточного ядра, заднее промежуточное ядро и латеральную часть переднего промежуточного ядра соответственно. Латеральная зона представлена областями D1 и D2, волокна которых адресуются мелко- и крупноклеточной частям латерального ядра соответственно.
Судя по имеющимся данным зоны А к Вмлекопитающих можно рассматривать как соответствующие медиальной и основному объему промежуточной зон рептилий; зона С - небольшому участку промежуточной зоны, локализованному у разных рептилий неодинаково (ростро-латерально у черепах и каудолатерально у варанов); флоккулус млекопитающих - латеральной зоне рептилий.
Следует отметить, что мозжечок млекопитающих традиционно подразделяли на древнюю часть архицеребеллум, старую часть - палеоцеребеллум и новый мозжечок, или неоцеребеллум. При этом исходили не только из характера распределения афферентов в пределах коры, но и из предполагаемого филогенетического развития. Основные связи, распределенные по этим отделам, позволили говорить о вестибуло-, спино- и понтоцеребеллярных отделах. Вероятно, подобное деление можно обнаружить и у птиц, у которых под неоцеребеллярной частью можно понимать зону окончания "мостовых входов" (латеральные отделы долек V-VIII). В мозжечке рептилий аналогичный подход позволяет выделить лишь старый мозжечок (спиноцеребеллум), объединяющий переднюю и среднюю доли, и древний (вестибулоцеребеллум), занимающий заднюю долю.
Вместе с тем следует иметь в виду, что подобное деление нельзя рассматривать как исчерпывающее, поскольку современные данные свидетельствуют о более сложном характере распределения входов в пределах мозжечка. Так, вестибулярные проекции у млекопитающих не ограничены флоккуло-нодулярной долей и оказываются рассеянными по большой поверхности полушарий. Мостовые входы не ограничены латеральными отделами полушарий, но включают и область червя. Наконец, при выделении неоцеребеллярной части мозжечка млекопитающих подчеркивают существование не только мостовых, но и оливарных и катехоламинергических входов в эту область, однако источники двух последних (нижняя олива и голубое пятно) являются прецеребеллярными структурами и у других позвоночных, в частности рептилий.
Эфференты мозжечка сформированы аксонами нейронов мозжечковых ядер и в некоторых случаях - клеток Пуркинье (когда речь идет о вестибулярных проекциях) (рис. 51). Ядра мозжечка помимо закономерно проецирующихся на них клеток Пуркинье получают коллатерали мшистых и лиановидных волокон, а также образуют небольшое количество проекций к ко-:ре мозжечка. Однако основная часть эфферентов направляется
за пределы мозжечка, к экстрацеребеллярным структурам в виде двух контрлатеральных трактов: соединительной ножки, bгаchium conjunctivum, и крючковидного пучка, fasc. uncinatus. Первая совершает перекрест при вхождении в средний мозг, второй - в пределах мозжечка.
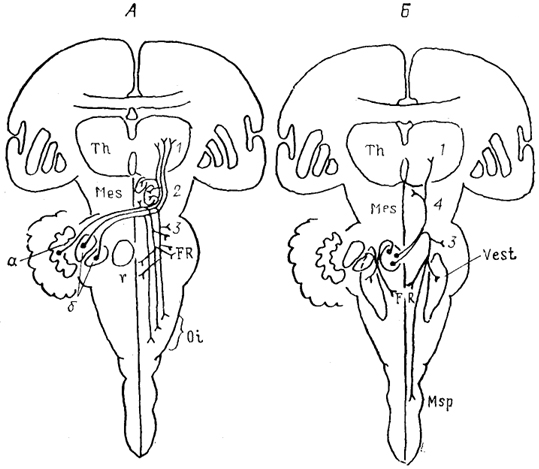
Рис. 51. Характер распределения эфферентов мозжечка приматов (Haines, 1986).
А - эфференты латерального (а) и промежуточных (б) ядер, Б - эфференты медиального ядра. 1- ядра таламуса, 2 - красное ядро, центральное серое вещество-среднего мозга и ядро III нерва, 3 - ядра моста, 4 - ядро задней комиссуры и передний холм.
Соединительная ножка берет начало от латерального ядра у птиц и рептилий и от промежуточного и латерального - у млекопитающих. У птиц и млекопитающих она формирует восходящую и нисходящую ветви, у рептилий - только восходящую. Восходящая ветвь у всех позвоночных адресована красному ядру, а у млекопитающих и птиц - и дорсальному таламусу (у рептилий аналогичные входы обнаружены только у некоторых ящериц и крокодилов). Вместе с тем для всех рептилий характерен прямой вход в теленцефалон (вместо опосредованного таламическими образованиями у птиц и млекопитающих). У млекопитающих часть волокон адресована ядрам конечного мозга. Нисходящая ветвь соединительной ножки направляется
к ретикулярной формации понтомедуллярного отдела и к нижней оливе (и к ядрам шва у млекопитающих).
Крючковидный пучок у птиц и рептилий сформирован нейронами обоих ядер, у млекопитающих - нейронами медиального ядра. Его волокна адресованы контрлатеральному вестибулярному комплексу, медиальной зоне ретикулярной формации и спинному мозгу. У млекопитающих имеется и восходящая ветвь, крючковидного пучка, волокна которой проходят в верхних ножках и распределяются контрлатерально в области центрального серого вещества среднего мозга, передних холмах, ядре задней комиссуры, таламусе и гипоталамусе (в других, чем соединительная ножка, отделах). Восходящая ветвь у рептилий не выделяется, однако у черепах и варанов отмечены входы крючковидного пучка в красное ядро.
Помимо названных, у млекопитающих описан еще и прямой фастигио-бульбарный тракт, tr. fastigiobulbaris, состоящий из двух компонентов. Первый - восходящий - берет начало от ростролатеральной части ядра шатра и проходит в верхних ножках вместе с ветвью крючковидного пучка. Точное место его окончания неясно. Второй - нисходящий - компонент отходит от ростральной части ядра шатра и, проходя в нижних ножках, адресуется вестибулярному комплексу, предлежащему ядру, ретикулярной формации.
Заключая обзор строения мозжечка позвоночных, следует подчеркнуть, что несмотря на разный уровень развития животных и их адаптивные особенности, этот отдел их головного мозга характеризуется целым рядом общих черт. В первую очередь это относится к нейронному строению и организации внутрицеребеллярных взаимодействий: у всех позвоночных наличие гранулярных нейронов и клеток Пуркинье, у высших - присутствие корзинчатых клеток, сходство возбуждающих и тормозных элементов. Применительно к характеру связей также можно выделить некоторые общие закономерности.
Среди источников афферентации у всех позвоночных имеются ядра октаво-латеральной системы, спинной мозг, нижняя олива, нисходящее ядро тройничного нерва, латеральная зона ретикулярной формации. Таким образом, консервативность афферентных связей прослеживается в отношении ромбовидного мозга. В то же время источники входов, расположенные на уровне среднего и переднего мозга, оказываются разными в разных группах. У птиц и млекопитающих - это опосредованные "мостовыми" ядрами входы от теленцефальных отделов, у рыб - претектальная область, у некоторых телеостей - паракомиссуральное ядро (ряд авторов полагает, что оно, как и у высших, обеспечивает передачу влияний от теленцефалона). Возможно, что ромбенцефаллические входы как наиболее древние действительно отличаются большей стабильностью и опосредуют организацию неких общих функций. Ростральные же отражают
как общий уровень развития мозга, так и адаптивные особенности (например, связи мозжечка с вышележащими отделами у электрических костистых рыб и у некоторых хрящевых). Эфферентные связи мозжечка также имеют общие черты: во всех группах обнаруживаются входы к октаво-латеральным центрам, ретикулярной формации, красному ядру (или его прекурсору). У высших позвоночных добавляются проекции в теленцефалон (прямо - у рептилий, или при посредстве дорсального таламуса - у птиц и млекопитающих). Общим для всех названных путей мозжечка является то, что они оказывают влияние на надсегментарные моторные образования, которые в свою очередь иннервируют моторные ядра головного и спинного мозга. В тех группах, где появляются высшие моторные системы, способные изменять активность надсегментарных и сегментарных моторных центров, влияния мозжечка адресуются и звеньям этих систем (моторная кора млекопитающих, мелкоклеточная часть красного ядра, стриатум). Таким образом, при незначительной выраженности прямых проекций мозжечка к спинному мозгу характер его связей обеспечивает возможность влияний на моторные системы любого уровня.
* Название "аурикулярные доли", таким образом, используются лишь для обозначения гомологичных структур при сравнительной характеристике. Предпочтительнее название "вестибулоцеребеллум" или "флоккулус".
* У некоторых черепах и.змей слой клеток Пуркинье не выражен, и последние диффузно распределены между молекулярным и гранулярным слоями.
* У млекопитающих коллатерали аксонов отходят в той же плоскости, что и дендриты.
* Эти входы либо непосредственно адресованы мозжечку (как коре, так и ядрам), либо поступают через прецеребеллярные ядра. В первом случае они формируют волокна, отличные и от мшистых и от лиановидных и получившие название "мультислойных", поскольку, проходя перпендикулярно через все слои коры, они отдают многочисленные коллатерали в гранулярном и ганглионарном слоях. Войдя в молекулярный слой, они разделяются и идут параллельно длинной оси долек. Точное место их окончания не ясно, однако они весьма сходны с КА-ергическими волокнами коры мозга.
ГЛАВА 5
СРЕДНИЙ МОЗГ
Средний мозг, Mesencephalon, является ростральным отделом ствола мозга позвоночных и в его состав входит продолжение сомато- и висцеросенсорных, а также сомато- и висцеромо-торных зон. Однако подобное разделение здесь не столь выражено, как в ромбовидном мозге, и составляющие этих колонн занимают лишь небольшой объем мезенцефального уровня. Основные же мезенцефальные структуры входят в состав моторных и интегративных систем мозга.
В составе среднего мозга выделяют производное крыловидной пластинки - крышу среднего мозга, tectum mesencephali, и производное базальной пластинки - покрышку, tegmentum mesencephali. У высших позвоночных (вследствие дальнейшего развития высших отделов мозга и их связей) возникает дополнительная часть - ножки мозга, pedunculi cerebri, - совокупность волокон теленцефального происхождения, адресованных эффекторным отделам ствола.
§ 1. СЕНСОРНЫЕ ЯДРА СРЕДНЕГО МОЗГА
Наиболее характерная для среднего мозга всех позвоночных структура - зрительный тектум, tectum opticum, у анамний представляет собой самую заметную часть мозга (см. рис. 5). Строение тектума подробно изучено у большого количества видов, принадлежащих разным классам. Во всех случаях типичным оказывается четко выраженное ламинарное строение данного образования. Число слоев, нейронный состав и организация связей в значительной степени варьируют, что определяется не только принадлежностью животных к разным таксонам, но и уровнем развития систем мозга, в первую очередь зрительной сенсорной системы, и организацией моторных взаимодействий.
Уже в мозге круглоротых тектум является крупным образованием, выдающимся латерально и ограничивающим выстланный эпендимой тектальный желудочек. Несколько типов и нейронов
с хорошо развитым дендритным полем, ориентированным радиально или тангенциально, формируют 6-8 страт, разделенных на слои. Основной объем афферентов поступает в наружные отделы тектума и проводит информацию от органов зрения; в более глубоких отделах обнаружены входы от других сенсорных образований - октаво-латерального комплекса и спинномозгового ядра тройничного нерва. Имеются сведения и о входах от мозжечка и спинного мозга, таламуса и эпиталамуса.
Эфференты тектума образуют несколько путей: текто-бульбарный (связывающий его с ромбовидным мозгом), текто-таламичеекий и текто-гипоталамический. Часть аксонов тектальных клеток распределяется в пределах тегментума среднего мозга. Таким образом, уже в мозге круглоротых в строении тектума и организации его связей прослеживаются черты, общие для всех позвоночных. Существенным является и характер распределения проекций: верхние отделы являются основной мишенью для ретинальных входов, эфференты же формируются в глубоких отделах тектума.
В мозге пластиножаберных тектум образован несколькими слоями, составленными нейронами разных типов. В дорсовентральном направлении они подразделяются на четыре страты: str. medullare externum, cellulare externum, medullare internum, cellulare internum. Верхние отделы принимают зрительные афференты, в то время как к двум нижним стратам подходят проекции от других отделов мозга - ретикулярной формации, теленцефальных структур, а также от контрлатерального тектума. На одном виде показано и существование соматосенсорных проекций. Эфференты, возникающие в тектуме, формируют восходящие, комиссуральные и нисходящие связи: восходящие адресованы претектальной области и зрительным ядрам таламуса; комиссуральные связывают одноименные структуры и интерколликулярное ядро; нисходящие объединяются под названием текто-бульбарного тракта и адресованы в основном структурам ретикулярной формации и интерколликулярному ядру. Несколько противоречиво решается вопрос о существовании у пластиножаберных текто-спинального тракта, свойственного практически всем позвоночным: одни авторы отрицают наличие у акул прямых связей тектума со спинным мозгом, другие отмечают его чрезвычайно слабое развитие. Связи тектума и ядер глазодвигательного комплекса, по-видимому, опосредуются ретикулярными образованиями. В целом тектум пластиножаберных имеет значительные черты сходства с другими позвоночными, однако некоторые авторы в числе особенностей данной структуры у пластиножаберных отмечают менее выраженную стратификацию, относительную бедность клеточных форм и как примитивную черту билатеральный характер теленцефальных входов. Отличительной особенностью тектума является также
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|