- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 7 страница
Описание комплекса ядер латеральной петли у птиц достаточно противоречиво. У видов с развитой слуховой системой большинство авторов выделяет в его составе верхнюю оливу, вентральную часть ядер латеральной петли и вентральное ядро латеральной петли. К каждому из этих отделов идут проекции от определенных участков ядер первого уровня, а их эфференты адресованы среднемозговому акустическому центру. Кроме того, вентральное ядро латеральной петли дает начало нисходящим проекциям к моторным ядрам, осуществляющим контроль за голосовым аппаратом, т. е. опосредует видоспецифические вокализации. От верхней оливы начинается путь, обеспечивающий эфферентную иннервацию улитки. Особенностью, не получившей достаточных объяснений, является существование прямых восходящих путей к теленцефалону, источник которых ряд авторов обозначает как промежуточное ядро латеральной петли. Сложность идентификации комплекса латеральной петли у птиц, по-видимому, обусловлена его тесными связями с окружающей ретикулярной формацией, что не позволяет очертить границы некоторых участков конкретных ядер.**
В пределах класса млекопитающих имеются значительные экологически обусловленные адаптации слуховой системы и, соответственно, наблюдается разная степень ее развития. Центральные отделы слуховой системы оказываются весьма развитыми
уже в понтомедуллярном отделе, где располагается .комплекс кохлеарных (улитковых) ядер, подразделяемый у разных животных на два и более самостоятельных отдела. Весьма сложно организованы и верхнеоливарный комплекс, трапециевидное тело, комплекс латеральной петли, состоящие из нескольких ядер. От каждого из названных отделов берут начало восходящие проекции к среднему и промежуточному мозгу, многочисленные связи с ретикулярными отделами и моторными ядрами ствола, а также обратные связи в пределах сенсорного канала.
У однопроходных и сумчатых в акустической области продолговатого мозга описаны крупноклеточное и угловое ядра, nucll. magnocellularis et angularis. У однопроходных они подобны одноименным образованиям мозга рептилий как по характеру связей, так и по типам нейронов. По расположению же, в частности относительно веревчатых тел, они оказываются сходными и у сумчатых и не только с ядрами рептилий, но и птиц. У плацентарных эти структуры располагаются латерально относительно веревчатых тел и представлены передним вентральным кохлеарным ядром, nucl. cochlearis ventralis anterior, гомологичным крупноклеточному ядру рептилий, и дорсальным и задним вентральным кохлеарными ядрами, nucll. cochlearis dorsalis et ventralis posterior, гомологичными угловому ядру сумчатых. У млекопитающих происходит дальнейшая дифференцировка слуховых центров в области моста: усложняется комплекс верхней оливы, развиваются ядра трапециевидного тела, в системе латеральной петли выделяются дорсальное, промежуточное и вентральное ядра.* Все названные образования в мозге млекопитающих формируют многочисленные связи в пределах специфического сенсорного канала, а также проекции к другим отделам мезга (моторным ядрам черепно-мозговых нервов, ретикулярной формации, центральному серому веществу среднего мозга), обеспечивая тем самым организацию системных защитных реакций (в частности за счет активации мышц среднего уха) и видоспецифических вокализаций. От верхнеоливарного комплекса берут начало оливо-кохлеарные тракты, осуществляющие иннервацию рецепторных отделов.
Акустические центры млекопитающих отличаются от таковых у птиц и рептилий более разнообразным нейронным составом,
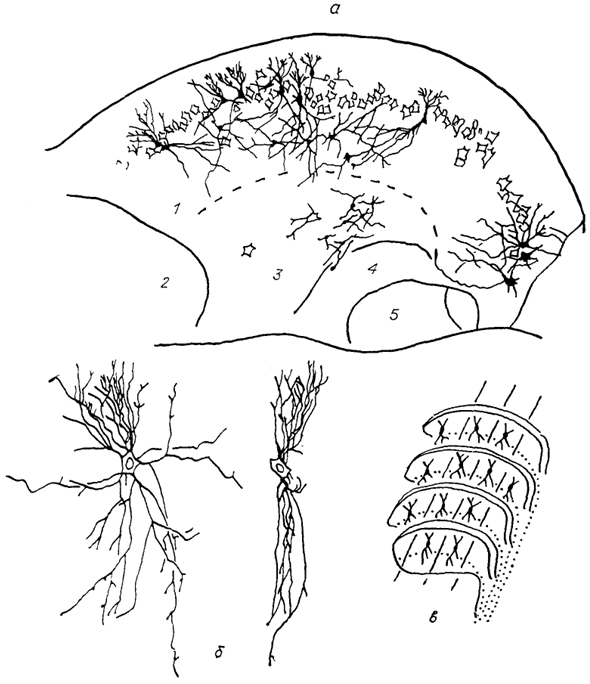
Рис. 40. Строение дорсального кохлеарного ядра кошки (Blackstadt e. а., 1984).
а - нейронный состав средней части дорсального кохлеарного ядра, б - пирамидоподобные нейроны, в - расположение пирамидоподобных клеток в ядре. 1 - слой гранулярных клеток, 2 - антеровентральное кохлеарное ядро, 3 - центральное ядро дорсального кохлеарного ядра, 4 - акустическая полоска, 5 - постеровентральное кохлеарное ядро.
многокомпонентностью и богатством связей внутри комплекса кохлеарных ядер и верхней оливы.
Среди кохлеарных ядер самым вариабельным оказывается дорсальное ядро. Оно наиболее развито у животных, ориентированных на восприятие звуков окружающей среды (хищные, зайцеобразные, грызуны), и имеет в этом случае четкое ламинарное строение (рис. 40). Напротив, в мозге эхолоцирующих млекопитающих, активность которых в значительной степени
обусловлена восприятием эха собственных сигналов, размеры дорсального ядра малы. Структурной особенностью этого ядра является наличие нейронов особой формы, напоминающих по ряду признаков клетки Пуркинье мозжечка. Они описаны под разными названиями (пирамидные, крупные биполяры, фузиформные) и располагаются в верхних слоях дорсального ядра перпендикулярно поверхности. Их дендриты ветвятся в одной, плоскости парасагиттально и пронизаны входящими под прямым углом афферентами. Аксоны этих клеток формируют слуховую полоску и проецируются на контрлатеральный задний холм. Другая особенность дорсального ядра - наличие разномодальных входов: помимо кохлеарных афферентов оно, в отличие от других улитковых ядер, получает проекции от главного ядра V нерва, вестибулярного комплекса и ядер дорсальных канатиков. Эфференты дорсального ядра направляются билатерально к верхнеоливарному комплексу, контрлатеральным ядрам латеральной петли и центральному ядру заднего холма. Ипсилатеральные проекции адресованы ретикулярной формации, медиальному и латеральному вестибулярным ядрам; и, вероятно, мозжечку.
Вентральные кохлеарные ядра проецируются на разные отделы верхнеоливарного комплекса. Кроме того, заднее вентральное ядро образует входы к моторному ядру V нерва ('иннервация небной занавески), двойному ядру (иннервация голосового аппарата) и ретикулярной формации. Эфференты вентрального ядра формируют трапециевидное тело, дорсального - слуховые полоски, corpus trapezoideum et striae acustica, соответственно. У некоторых млекопитающих показаны комиссуральные связи кохлеарного комплекса.
Верхнеоливарный комплекс млекопитающих значительно дифференцирован и имеет в качестве основных три отдела: медиальную верхнюю оливу, oliva superior medialis, латеральную верхнюю оливу, oliva superior lateralis, медиальное ядро трапециевидного тела, nucl. medialis corporis trapezoidei (рис. 41). У представителей разных отрядов степень их развития различна. Так, у приматов наблюдается постепенное увеличение размеров медиальной верхней оливы, у человека она достигает максимального развития. Латеральная верхняя олива и ядро трапециевидного тела велики у рукокрылых, насекомоядных, китообразных. У хищных и грызунов хорошо представлены все отделы комплекса.
Эфференты каждого из компонентов верхнеоливарного комплекса направляются к разным отделам вышележащих слуховых структур: ядрам латеральной петли и задним холмам. У некоторых видов описаны прямые проекции в таламус. Другие связи идут к моторным ядрам черепномозговых нервов (обеспечивая регуляцию вспомогательных структур периферического отдела слуховой системы), ретикулярной формации,
кохлеарным ядрам. Наконец, с этого уровня у млекопитающих осуществляется иннервация внутренних и наружных волосковых клеток волокнами ипси- и контрлатерального оливо-кохлеарных трактов соответственно.
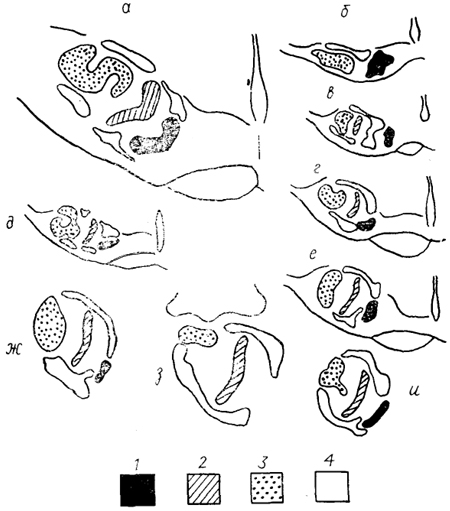
Рис. 41. Верхнеоливарный комплекс млекопитающих (Moore, 1987).
а - кошка, б - еж, в - землеройка, г - мартышка, д - галаго, е - совинолицая мартышка, ж - макака, з - человек, и - гиббон. 1 - трапециевидное тело; 2, 3 - медиальная (2) и латеральная (3) верхние оливы; 4 - периоливарные ядра.
Дорсальное ядро латеральной петли у млекопитающих характеризуется относительной стабильностью развития в мозге плацентарных. Оно получает входы от контрлатерального кохлеарного комплекса, ипсилатеральных медиальной верхней оливы и вентрального ядра латеральной петли, а также билатеральной верхней оливы. Эфференты дорсального ядра в основном восходят билатерально к задним холмам, образуя латеральную петлю, lemniscus lateralis, а также проецируются на контрлатеральный верхнеоливарный комплекс. Вентральное ядро латеральной петли слабо выражено у приматов и человека и сложно организовано у эхолоцирующих животных. Основные входы оно получает от кохлеарных ядер, а его эфференты в основном направлены к ретикулярной формации (что объясняет участие этой структуры в организации такой реакции, как startl-reflex), часть проекций восходит к задним холлмам.

Рис. 42. Схема преобразований акустико-латеральной системы позвоночных (Fritzsch, 1988).
1 - круглоротые (Lamvetra); 2-8 - рыбы: Polypterus (2), Squalus (3), Amta (4), Salmo (5), Latlmeria (6), Lepldoslren (7), Siren (8); 9-15 амфибии: Salamandra (9), Trlturus (10), Bolitoglossa (11), Ichthyophis (12), Xenopus (13), Rana (14), Bombina (15); 16 - рептилии (Testudo); 17 - птицы (Anas); 18 - млекопитающие (Didelphis).
У некоторых млекопитающих хорошо развито и промежуточное ядро латеральной петли, связанное с системой вокализации и участвующее в так называемом центральном ослаблении акустических сигналов у эхолоцирующих животных.
Отметим, что у всех амниот организация связей слуховой системы оказывается сходной. Так, центры первого и второго порядков (кохлеарные ядра, комплекс верхней оливы) формируют восходящий путь, адресованный среднему мозгу. Нейроны мезенцефалического уровня проецируются на дорсальный таламус, а через него на теленцефалои: в паллиальные (или стриатарные - у птиц) отделы. В процессе эволюции слуховой системы происходит усиление бинауральных взаимодействий, появление "спрямленных" восходящих и нисходящих путей.
Все ядерные формации слуховой системы амниот рассматриваются
как гомологичные, в то время как гомологизация их с подобной системой низших затруднена: убедительные доказательства единства происхождения слуховых центров мозга амниот и амфибий отсутствуют. Вопросы гомологизации не решены и в пределах группы рыб и между рыбами и амфибиями. Возможно, следует согласиться с авторами, допускающими независимость возникновения и параллельное развитие слуховых центров, по крайней мере, у амфибий и амниот. Что же касается рыб, то учитывая гетеромодальность рецепторов октаво-латеральной системы, трудно рассчитывать на обнаружение какой-либо предковой формы, общей для всех этих животных и давшей начало развитию акустической системы в разных группах (рис. 42).
Тем не менее не следует забывать, что между всеми названными системами - органов боковой линии, вестибулярной и слуховой - можно обнаружить значительное сходство. Оно касается и рецепторов, и путей проведения в мозге, и наличия специфических отделов, обеспечивающих эфферентный контроль рецепторных образований. Наконец, для акустической системы и системы органов боковой линии показано существование уровней, в которых обнаруживается полное представительство рецептивной поверхности и создаются "карты пространства" (в торусе и верхней оливе и в тектуме соответственно).
* У низших позвоночных информация от соматических рецепторов передается не только от тройничного ганглия (по системе тройничного нерва), но и от сенсорных ганглиев VII, IX и X черепномозговых нервов. Однако у всех челюстноротых гипертрофия мандибулярной дуги отражается в доминировании сенсорного входа именно через V нерв (краниальный нерв мандибулярной дуги).
* Комплекс этих черепномозговых нервов у низших позвоночных отчасти входит в состав соматосенсорной системы. Однако даже у рыб соматический компонент этих нервов иннервирует внешние покровы лишь в области наружных жаберных щелей. У четвероногих происходит дальнейшее уменьшение объема соматического компонента, а у млекопитающих единственной зоной, иннервация которой не обеспечивается системой тройничного нерва, является область наружного слухового прохода, иннервируемая клетками яремного ганглия.
* Отметим, что волокна вестибуло-кохлеарного нерва адресованы и мозжечку.
** Крупноклеточное вестибулярное ядро получает также входы нервов боковой линии.
* Входящее в состав октаво-латеральной области амфибий дорсолатеральное ядро рассматривается как слуховое, и его описание будет дано ниже.
* Для костистых рыб получающими акустическую информацию считаются тангенциальное ядро и ядро Дейтерса. У скатов Platyrhinoidis активность в ответ на предъявление звуковых стимулов зарегистрирована в ядрах октавного комплекса, и латеральной петле.
* У некоторых рептилий обнаружены комиссуральные связи между ядрами кохлеарного комплекса.
* Этим обстоятельством можно объяснить противоречивость сведений о комплексе латеральной петли птиц, включая утверждение о гомологии вентрального ядра птиц парабрахиальному комплексу грызунов, а дорсального - мезенцефалической ретикулярной формации.
* При этом остается неясным вопрос об их гомологах в мозге птиц. Так, принято рассматривать медиальную верхнюю оливу как соответствующую ламинарному ядру, дорсальное ядро латеральной петли как гомолог вентральной части ядер латеральной петли птиц, а вентральное ядро латеральной петли млекопитающих - одноименному ядру птиц. Что же касается латеральной верхней оливы, медиального ядра трапециевидного тела и промежуточного ядра латеральной петли - ядер хорошо развитых у многих млекопитающих - то их гомологизация с мозгом птиц не проведена.
§ 3. МОТОРНЫЕ ЯДРА ПРОДОЛГОВАТОГО МОЗГА
Висцеромоторная зона занимает вентролатеральные отделы продолговатого мозга и содержит ядра V (тройничного), VII (лицевого), IX (языкоглоточного) и X (блуждающего) нервов, опосредующих специализированные бранхиомоторные реакции. У круглоротых и рыб ядра этой зоны не формируют дифференцированных агрегаций и в ряде случаев трудноотделимы от их сенсорных звеньев.* Тем не менее, если у голостей и хрящевых рыб ядра этой зоны занимают перивентрикулярную позицию (рис. 34), то у большинства телеостей моторные ядра V и VII нервов мигрируют от стенки желудочка вглубь и оказываются отделенными от других ядер (рис. 28, 35). Во многих отношениях развитие указанного комплекса коррелирует с объемом вторичного вкусового тракта.
Нейроны ядер V и VII нервов иннервируют шесть наиболее важных дыхательных мышц. При этом клетки, иннервирующие мышцы, которые участвуют в разных фазах дыхательного цикла, локализованы в различных областях ядер. Основные афференты к данному комплексу образованы аксонами нейронов
нисходящего ядра V и сенсорного ядра VII нервов. Другими входами служат проекции ретикулярных образований, главного ядра V нерва, заслонки мозжечка, а также некоторых ядер промежуточного мозга.
У амфибий и рептилий моторное ядро V нерва представляет собой самостоятельный нейронный комплекс (рис. 30, 31), его развитие, как следует из сравнительноанатомического анализа, коррелирует со сложностью организации максиллярной и небной мускулатуры, в частности, оно велико у ящериц и змей. У этих животных ядро V нерва состоит из трех компонентов и образовано несколькими типами клеток, сходных с описанными у амфибий и птиц. Однако гомологизация этих отделов затруднена, поскольку у птиц моторное ядро тройничного нерва объединено с ядром лицевого нерва, которое у рептилий образует единый комплекс с ядром IX нерва. Тем не менее, полагают, что один тип нейронов у рептилий напоминает элементы, образующие у млекопитающих добавочные моторные ядра V и VII нервов и иннервирующие мышцы основания ротовой полости. Доказательств же раздельной иннервации разных мышечных групп на рептилиях не получено. Входы к данному ядру сформированы вестибулярным комплексом, мозжечком, а также сенсорными ядрами ствола (в основном тригеминальным комплексом).
У высших позвоночных основные входы к моторному ядру V нерва образуют главное и нисходящее ядра и ретикулярная формация. У высших млекопитающих имеются кортикальные проекции, которые у низших (например, у крыс) отсутствуют. Эфференты направляются к мышцам, открывающим рот и сжимающим челюсти.
Другие ядра висцеромоторной зоны представлены в мозге позвоночных различным образом. У хрящевых рыб ядра VII, IX и X нервов образованы колонной крупных нейронов, иногда разделенных волокнами. В ее каудальной части, в области, соответствующей ядру X нерва, размеры клеток несколько меньше, и эту область ряд авторов обозначает как медиальное моторное ядро блуждающего нерва, nucl. motorius medialis n. vagi, гомологизируя его с дорсальным моторным ядром X нерва высших позвоночных. Латерально от общей моторной колонны располагается латеральное моторное ядро X нерва, nucl. motorius lateralis n. vagi, - предполагаемый гомолог двойного ядра высших. Однако недостаточность сведений о характере его связей и особенностях нейронной организации не позволяет утверждать это однозначно: если у рыб размеры нейронов латерального ядра меньше, чем в медиальном ядре, то у высших позвоночных нейроны двойного ядра по размерам превышают таковые в дорсальном моторном ядре.
Примером модификации структур ромбовидного мозга служат изменения его внешнего вида и формы у электрических скатов, электрические органы
которых возникают за счет преобразований плавников. Мотонейроны, регулирующие активность этих органов, сосредоточены в особых электрических долях (по некоторым данным они составляют до 60% массы мозга), насчитывающих 120 тыс. мультиполярных клеток. Аксоны электромотонейронов формируют четыре электрических нерва, иннервирующих электрические органы. Происхождение этих нервов до конца не выяснено, чаще всего их относят к системам VII, IX и X нервов.
У актиноптеригий и амфибий единая клеточная группа нейронов IX и X нервов, по-видимому, является предшественником двойного ядра, а моторная часть VII нерва представлена самостоятельной группой клеток, лежащих под сенсорным ядром.
Описания этого комплекса у рептилий весьма противоречивы. Так, у ящериц областью иннервации VII и IX нервов являются различные мышцы головы, причем некоторые из них имеют двойную иннервацию, что может быть следствием их происхождения (из 2-й и 3-й бранхиальных дуг). В пределах ядер показана топологическая организация нейронов, иннервирующих разные мышцы. Вместе с нейронами ядра V нерва эти клетки составляют основную часть вентролатеральной колонны продолговатого мозга ящериц. Моторное ядро VII нерва связано с иннервацией нескольких мышечных групп, и крупные нейроны этого отдела образуют несколько подъядер. Однако в ряде случаев его элементы объединены с клетками ядра IX нерва.
Большинство авторов полагает, что именно у рептилий происходит формирование двойного ядра, nucl. ambiguus, представленного в мозге высших позвоночных и являющегося объединением моторных элементов IX и X нервов. В мозге многих рептилий оно составлено относительно крупными нейронами и не имеет еще четких границ с другим ядром, характерным для этой области высших, - дорсальным моторным ядром
X нерва. Отметим, что двойное ядро рептилий (в том смысле, в каком его понимают у высших, т. е. совокупность нейронов, иннервирующих гортань), представлено лишь клетками блуждающего нерва. Клеточный состав его такой же, как и у других ядер комплекса. Мышцы, которые им иннервируются, происходят из 4-й бранхиальной дуги и формируют мышечную стенку глотки, а также специальную мышцу - констриктор внутренней яремной вены.
У птиц афференты двойного ядра поступают от сенсорных ядер вышележащих отделов мозга, включая гипоталамус и конечный мозг, а также области, входящей в состав комплекса латеральной петли и связанной с осуществлением вокализаций. Возможно, что конвергенция входов от разных источников обеспечивает именно с этого уровня сочетанный контроль дыхания и звуковоспроизведения.
Моторное ядро VII нерва млекопитающих получает многочисленные входы от мезенцефалического и понтомедуллярного
отделов. Кроме того, через красное ядро сюда поступают проекции от новой коры. Мотонейроны ядра иннервируют поверхностные мышцы головы и шеи, а также стремечковую мышцу. Благодаря наличию разнообразных источников иннервации это ядро принимает участие в осуществлении многих приспособительных реакций. Так, входы от паралемнисковой области в медиальную часть ядра обеспечивают ориентацию ушных раковин в сторону источника звука, входы от центрального серого вещества среднего мозга - вокализации, а проекции от ядер Колликера-Фузе и вентрального парабрахиального - такие реакции, как принюхивание. У хищных обнаружено небольшое количество входов от вестибулярного комплекса.
Размеры моторного ядра лицевого нерва у млекопитающих соответствуют степени развития мускулатуры головы. Оно небольшое у утконосов, сложно дифференцировано у слонов и достигает значительной выраженности у человекообразных обезьян в связи с развитием мимической мускулатуры.
Двойное ядро млекопитающих представлено совокупностью крупных мультйполяров, иннервирующих ларингеальные и фарингеальные мышцы. У многих млекопитающих оно меньше, чем моторное ядро VII нерва. Максимального развития достигает у эхолоцирующих рукокрылых и китообразных. Среди животных, не обладающих этой способностью, оно наиболее развито у некоторых представителей ленивцев. Основные проекции к двойному ядру поступают от структур вкусовой сенсорной системы (от ядра солитарного тракта, парабрахиальных ядер) от центрального серого вещества среднего мозга, от новой коры. В случае значительного развития оно непосредственно переходит в ядро лицевого нерва, и у некоторых животных в его состав включают рострально и каудально лежащие группы клеток - nucll. retrofacialis et retroambiguus соответственно. В пределах ядра, а также в непосредственной близости от него обнаружены парасимпатические нейроны, осуществляющие респираторный контроль. Такие клетки сосредоточены в ретро-фациальной части, а также в вентролатеральном отделе ядра, в области, получившей название периамбигуальной.
В целом между моторными ядрами существуют достаточно-тесные взаимосвязи (прямые или, чаще, через ретикулярную формацию), что обеспечивает их участие в организации системных реакций.
Комплекс висцеромоторных ядер содержит и преганглионарные элементы вегетативной нервной системы, однако для низших позвоночных их систематизированное описание отсутствует. Так, у хрящевых рыб источником преганглионарных волокон служит область ядра X нерва, но функциональная роль этих связей не выяснена. Для актиноптеригий описаны многочисленные парасимпатические ветви, берущие начало от ядра X нерва. Точная локализация их источников пока не известна,
но вероятно, данные нейроны составляют группу клеток, лежащую латерально от бранхиальных отделов этого комплекса (ядра VII, IX, X нервов), и таким образом представляют собой гомолог дорсального моторного ядра блуждающего нерва у высших.
У амфибий группа преганглионарных нейронов, по-видимому, сосредоточена в непосредственной близости от висцеромоторного комплекса и рассматривается как примордиальное моторное дорсальное ядро X нерва.
У рептилий, как полагают, происходит формирование дорсального моторного ядра X нерва, nucl. motorius dorsalis n. vagi, как самостоятельной части в результате разделения центрального представительства моторных элементов блуждающего нерва. У многих представителей клетки, которые обеспечивают иннервацию висцеральных органов, располагаются в дорсальном отделе нейронного комплекса. У черепах висцеромоторная часть X нерва четко отделена от бранхиомоторной. Следует отметить, тем не менее, что во всех группах, где проводился анализ структуры дорсального моторного ядра X нерва, показано наличие в нем компонентов VII и IX нервов.
У некоторых рептилий нейроны, дающие начало преганглионарным волокнам, обнаружены и в мелкоклеточной части ретикулярной формации, где они сосредоточены между ядрами солитарного тракта и лицевого керва, а также между двойным ядром и дорсальным моторным ядром X нерва. По расположению они соответствуют парасимпатическим ядрам млекопитающих и птиц, однако степень концентрации клеток в этих отделах мозга рептилий невелика даже у ящериц, у которых они наиболее выражены.
Дорсальное моторное ядро млекопитающих и птиц - протяженная структура, у некоторых (например, крыс) доходящая до шейных сегментов спинного мозга. Основные проекции к нему образованы аксонами нейронов ядра солитарного тракта, тригеминального комплекса, гипоталамуса. Эфференты направляются к многочисленным парасимпатическим ганглиям.
На уровне продолговатого мозга выделяется также группа клеток, лежащая между бранхиомоторными ядрами и связанная с регуляцией слезных и слюнных желез - верхнее и нижнее слюноотделительные ядра, nucll. salivatorius superior et inferior, принадлежащие системам VII и IX нервов. Ряд нейронов сосредоточен в пределах ретикулярной формации - периамбигуальная область и парвоцеллюлярное ретикулярное ядро - и связан с кардиореспираторной регуляцией.
В составе висцеромоторной колонны тетрапод выделяют комплекс нейронов, образующих ядро добавочного черепномозгового нерва, nucl. n. accessorii. Полагают, что он состоит из бульбарного и спинального компонентов. У млекопитающих первый иннервирует мышцы гортани и происходит из каудальной
части двойного ядра (именно на этом основании XI нерв относят к висцеромоторным). Спинальный компонент, вероятно, происходит из нескольких других ядер, включая дорсальное моторное ядро X нерва и моторные ядра шейных сегментов спинного мозга. Сфера иннервации этого компонента - мышцы, обеспечивающие движения головы.
Вопрос о происхождении XI нерва еще не решен окончательно. Уже у амфибий, как подчеркивают некоторые авторы, имеется разделение центрального представительства X и XI нервов и различие в форме и размерах клеток. Данные, касающиеся рептилий, противоречивы. Так, в ряде работ в составе XI нерва обнаружены корешки X нерва, что подтверждает его гомологичность бульварному компоненту млекопитающих. Другие результаты противоречат этому: нейроны ядра XI нерва у рептилий оказываются в 2-3 раза превосходящими по размеру клетки двойного ядра и в 5 раз - нейроны дорсального моторного ядра X нерва. Эти данные, а также пространственное разделение ядер позволили ряду авторов отстаивать самостоятельное происхождение добавочного нерва. Сферой его иннервации являются мышцы, образующиеся из вентральной части мезенхимы 4-й и, возможно, 5-й бранхиальных дуг (трапециевидная и грудино-ключично-сосцевидная и их. гомологи у других животных). Они иннервируются наиболее каудальной частью бульбарных мотонейронов, которые, как полагают, мигрировали в шейные сегменты спинного мозга. Исходя из этих данных XI нерв следует считать отдельным краниальным нервом, контролирующим движения головы.
Соматомоторная зона продолговатого мозга рассматривается как ростральное продолжение передних рогов спинного мозга. В ней располагаются ядра III, IV, VI и XII черепномозговых нервов, а у рыб, лишенных XII нерва, - центральное представительство так называемых спинно-окципитальных нервов (рис. 26).* В ее пределах расположены также крупноклеточные области медиальной ретикулярной формации и проходит медиальный продольный пучок, fasc. longitudinalis media-lis, волокна которого связывают различные образования ствола со спинным мозгом. Следует отметить, что в области, соответствующей соматомоторной зоне, расположены и такие структуры, как нижняя олива и дугообразные ядра (медуллярный отдел) и межножкозое ядро (мезенцефалический отдел) - образования, не являющиеся моторными, но имеющие отношение к моторным системам мозга.
В пределах продолговатого мозга располагаются центральные проекции двух черепномозговых нервов - подъязычного (XII), nucl. n. hypoglossi, и отводящего (VI), nucl. n. abducentis. Ядро VI нерва вместе с ядрами III и IV -нервов составляют единый комплекс ядер глазодвигательных нервов (он будет описан в гл. 5).
В мозге хрящевых рыб и актиноптеригий спинальная моторная колонна продолжается в область продолговатого мозга и составлена крупными мультиполярами. Эфференты этого отдела выходят в виде одного или более спинно-окципитальных нервов.
У амфибий в составе спиномоторной колонны выделяются крупные и средних размеров нейроны с сильно развитым дендритным полем. Ростральная ее часть отделена от остальной и рассматривается как примордиальное ядро XII нерва (у аксолотля оно не выделяется как самостоятельное). У других амфибий ядро подъязычного нерва состоит из дорсомедиального и вентролатерального компонентов, крупные нейроны которых дают начало эфферентным корешкам.
У многих рептилий ядро XII нерва не является сложным и представляет собой единую клеточную массу, лежащую вблизи медиального продольного пучка, но отделенную от него ретикулярной формацией (рис. 33), а у змей - и слабо отделенную от передних рогов спинного мозга. Однако, у некоторых рептилий ядро XII нерва является самостоятельным образованием и содержит несколько клеточных групп. Так, для черепах и крокодилов описаны не только главное, но и добавочное ядра. Еще более сложно это ядро организовано у хамелеона.
В мозге птиц и млекопитающих ядро XII нерва хорошо выражено, при этом у млекопитающих развитие мускулатуры языка приводит к увеличению объема и усложнению его центральных проекций. Так, для многих хищных характерно наличие нескольких клеточных групп. Кроме входов от сенсорных стволовых образований, свойственных ядру XII нерва всех позвоночных, у млекопитающих имеются входы от кортикальных областей, и именно они составляют основной объем афферентов данного ядра.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|