- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 9 страница
Медиальная часть состоит из молекулярного, ганглионарного и гранулярного слоев. Гранулярные клетки в основном образуют крупное возвышение - медиальную гранулярную область. Медиальная и латеральные части вестибуло-латеральной области мозжечка скатов соединены интераурикулярной гранулярной полоской.
Связи мозжечка пластиножаберных, особенно его тела, изучены далеко не в полной мере. У некоторых акул обнаруживаются проекции мезенцефалического ядра тройничного нерва, адресованные передней части тела мозжечка, точнее его клеткам Пуркинье. Спинномозговые проекции формируют билатеральные ростральный и каудальный тракты, trr. spinocerebellaris rostralis et caudalis,* волокна которых в основном распределяются в средней части тела мозжечка в виде типичных мшистых волокон, терминирующих на нейронах гранулярного возвышения и, в меньшей степени, на ядрах мозжечка. Вместе со спинальными волокнами в мозжечок поступают эфференты нижней оливы. Как и у высших позвоночных, их называют лиановидными волокнами, однако оканчиваются они на соме и проксимальных участках дендритов клеток Пуркинье. У скатов обнаружены также входы от претектальной области.**
Эфферентными элементами тела мозжечка являются клетки Пуркинье, аксоны которых направляются к ядру мозжечка. Следует подчеркнуть, что аксоны клеток Пуркинье тела мозжечка пластиножаберных не покидают пределы мозжечка и не образуют его эфферентов.
Связи вестибулярного отдела мозжечка (аурикул, или вестибуло-латеральной доли) изучены более подробно. Так, получило экспериментальное подтверждение представление о наличии сенсорных афферентов - волокон VIII нерва и нервов боковой
линии. При этом вестибулярные входы сосредоточены в основном в медиальной части аурикул, от органов боковой линии - в верхнем листке аурикул (у акул) или в латеральной гранулярной области (у скатов). Помимо первичных сенсорных имеются и афференты от октаво-латеральной области мозга и спинальные входы. Дорсальный гранулярный гребень скатов лишен афферентов от первичных и вторичных центров октаво-латеральной системы, основные входы в эту область сформированы тегментумом среднего, ромбовидного и спинного мозга. Эфференты латеральной части сходны: и от латеральной гранулярной области, и от дорсального гранулярного гребня основные проекции направляются к медиальной части и телу мозжечка. Медиальная часть проецируется в виде мшистых волокон на тело мозжечка и в виде параллельных волокон - в мозжечковый гребень. Кроме того, отсюда берет начало церебелло-октавный тракт, tr. cerebellooctavus, осуществляющий регуляцию октавных ядер. Он образован аксонами клеток Пуркинье, которые, как показано на Scyliorhinus, оказывают тормозное влияние на нейроны октавного комплекса. Полагают также, что от медиальной части начинаются пути к комплексу глазодвигательных ядер.
Таким образом, вестибулоцеребеллум (аурикулы и вестибуло-латеральная доля) пластиножаберных в основном связан с октаво-латеральной областью продолговатого мозга и, по крайней мере у скатов, не проецируется непосредственно на ядро мозжечка, он либо образует самостоятельные эфференты, либо связан с телом мозжечка.
Эфференты мозжечка сконцентрированы в трех церебелло-фугальных системах. Мозжечково-вестибулярный и мозжечково-бульбарный тракты, trr. cerebellovestibularis et cerebellobulbaris, адресуются ретикулярной формации и октаво-латеральной области ромбовидного мозга и могут быть представлены аксонами как нейронов мозжечкового ядра, так и клеток Пуркинье вестибулоцеребеллума. Соединительная ножка мозжечка, brachium conjunctivum, - основной эфферентный путь, сформированный нейронами ядра мозжечка. Его волокна проходят в средний мозг и после перекреста образуют восходящую и нисходящую ветви. Первая содержит волокна, адресованные прекурсору красного ядра, ядрам III и IV черепномозговых нервов и, незначительное число входов, в промежуточный мозг. Нисходящая ветвь иннервирует нижнюю оливу и медиальную ретикулярную формацию. Часть волокон ножки мозжечка образует ипсилатеральные входы к ретикулярной формации среднего мозга. Церебелло-моторный тракт, адресованный моторным ядрам, проходит в составе медиального продольного пучка.
В целом мозжечок пластиножаберных является высокоорганизованным в функциональном отношении отделом мозга и в его строении проявляется значительное число признаков, характерных
для высших позвоночных. Иными словами, несмотря на некоторые примитивные черты (относительная бедность клеточными типами, отсутствие соматотопического характера распределения эфферентов) уровень развития мозжечка данной группы животных сопоставим с его развитием у некоторых млекопитающих. Принципиально важным в этом отношении оказываются, во-первых, сходство организации эфферентов: клетки Пуркинье → ядро мозжечка → экстрацеребеллярные структуры, во-вторых, наличие восходящих проекций к промежуточному мозгу, и в-третьих, приуроченность основных эфферентов к надсегментарным моторным зонам мозга - ретикулярной формации и "красному ядру". Сходство подчеркивается и тем обстоятельством, что мозжечок акул играет модулирующую роль в организации моторной активности так же, как у высших позвоночных. Следует отметить и наличие в мозжечке, по крайней мере у скатов, отдела, где конвергирует разномодальная афферентация и, возможно, присутствует опосредованный вход от конечного мозга - дорсальный гранулярный гребень.
У представителей костных рыб строение мозжечка еще более различается, и встречается несколько вариантов его анатомических преобразований. Так, у саркоптеригий мозжечок относительно прост и разделяется на тело и аурикулярные доли. Последние хорошо выражены у латимерии (что связывают с высокоразвитой системой органов боковой линии) и двоякодышащих. Тело мозжечка небольшое, выпуклое у латимерии, у большинства двоякодышащих скрыто под тектумом (рис. 5, 6, 7). В его составе можно выделить молекулярный, ганглионарный и гранулярный слои (хотя у двоякодышащих клетки Пуркинье не везде образуют четкий слой).
Среди представителей лучеперых рыб обнаружены группы, обладающие мозжечком необычной формы из-за инвагинации его тела в полость желудочка (многоперы и хондростеи). При этом у многоперов типичные клеточные элементы не формируют чередующихся слоев. Аурикулы у хондростеи хорошо выражены и подобны имеющимся у акул, а у многоперов малы (см. рис. 5, 9, 10).
Голостеи, так же как и многоперы, имеют эвертированный мозжечок с относительно большим телом. Каудальная его часть образует гребень, проходящий над IV желудочком, и состоит из гранулярных клеток, покрытых молекулярным слоем (см. рис. 5, 11).
У костистых рыб мозжечок может быть относительно мал (сельдеобразные и угри), но может достигать и огромных размеров (клюворылообразные) (рис.44).
В целом же у лучеперых мозжечок состоит из тела, заслонки и вестибуло-латеральных долей.
Заслонка является наиболее вариабельной частью мозжечка
по сравнению с другими отделами.* Очень велик разброс Б степени ее развития у костистых рыб. У пескаря и морского черта это небольшой участок мозжечка, лежащий под тектумом. У карповых и сомообразных заслонка, напротив, столь развита, что вызывает изменение в конфигурации мозга, смещая каудальные отделы тектума латерально. Наконец, у клюворылообразных рода Mormyrus на долю мозжечка приходится 55% массы мозга, причем в основном за счет необычно крупной заслонки, покрывающей дорсально головной мозг (рис. 45).
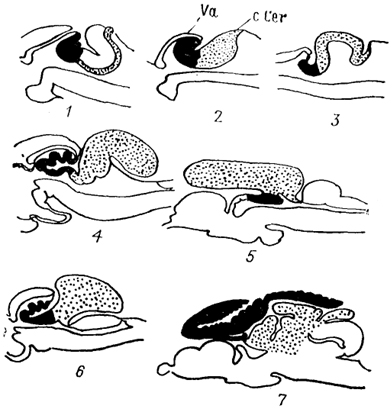
Рис. 44. Строение мозжечка костистых рыб (Nieuwenhuys, 1982).
1 - Erpetoichthys, 2 - Scaphirhynchus, 3 - Amia, 4 - Salmo, 5 - Glorias, 6 - Xenomystus, 7 - Gnathonemus.
В основании мозжечка рыб выделяется . группа клеток, составляющая ядро мозжечка, nucl. lateralis cerebelli. У хондростей и голостей оно представлено диффузно лежащими мультиполярными нейронами на границе ромбовидного мозга и мозжечка. У костистых рыб ядро четко оформлено, а в дорсальной части тегментума среднего мозга описана группа мелких, плотно лежащих клеток, составляющих ядро заслонки, nucl. lateralis valvulae cerebelli.
Тело мозжечка в общих чертах сходно построено у всех лучеперых рыб и содержит набор одинаковых элементов (развитый в разной степени), образующих гранулярный, ганглионарный и молекулярный слои. У костистых рыб в ганглионарном слое кроме типичных клеток Пуркинье описаны так называемые эвридендроидные клетки - крупные элементы с аксоном большого диаметра, выходящим за пределы мозжечка.
Вестибуло-латеральная доля у лучеперых представлена по-разному, и у хондростей и голостей напоминает аурикулы акул. У костистых рыб она не столь выражена и образована каудальной долей, lobus caudalis (поперечная пластинка, отделенная от тела мозжечка задней бороздой, fissura posterior), и парными
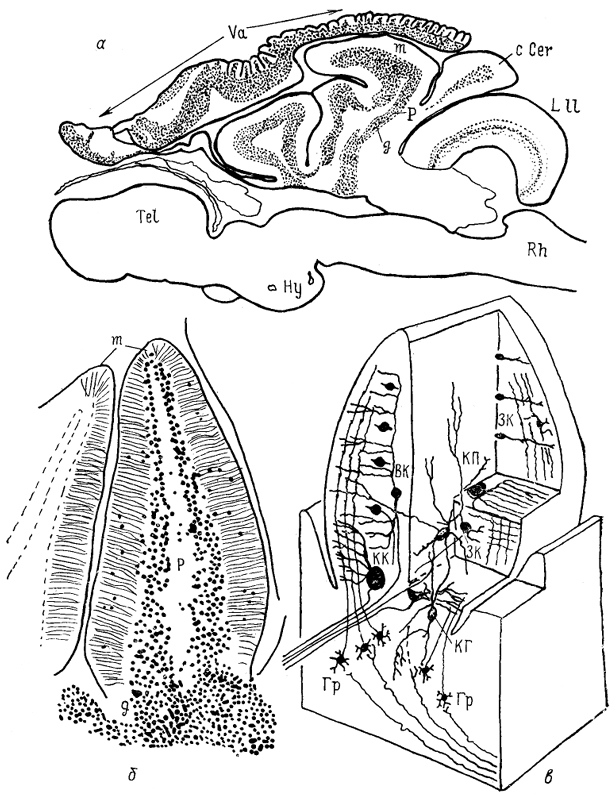
Рис. 45. Строение мозжечка представителя мормирид Gnathonemus petersi (Nieuwenhuys, 1969).
а - общий вид мозга на сагиттальном срезе; б, в - общий вид (б) и нейронный состав (в) дольки мозжечка. ВК - вертикальная клетка, ГР - гранулярная, ЗК - звездчатая, КГ - клетка Гольджи, КК - корзинчатая, КП - клетка Пуркинье.
гранулярными возвышениями, eminentiae granulares. В составе каудальной доли в дорсовентральном направлении выделяют слой гранулярных клеток, переходящий в гранулярные возвышения, фибро-молекулярный слой и парное скопление гранулярных клеток в перивентрикулярной области.
Афференты мозжечка лучеперых происходят из разных источников. У всех групп данного подкласса обнаружены входы от октаво-латеральной области продолговатого мозга, распределяющиеся в теле и заслонке. Не менее характерны и оливо-мозжечковые проекции в виде лиановидных волокон (не имеющих, однако, свойственного высшим позвоночным вида), оканчивающихся на телах и проксимальных участках дендритов клеток Пуркинье. Кроме того, для представителей всех изученных групп показаны спинно-мозжечковые входы и проекции от латерального клиновидного ядра.
Для костистых рыб и голостей характерно наличие первичных сенсорных проекций систем VIII нерва и нервов боковой линии; их упорядоченное распределение обнаружено во всех отделах мозжечка. Афференты мезенцефалического происхождения сконцентрированы в двух трактах: переднем и заднем, trr. mesencephalocerebellaris anterior et posterior. Задний тракт хорошо выражен у голостей и телеостей и проводит в тело мозжечка информацию от ядра заслонки, торуса и тегментума. Передний тракт, связывающий тектум с гранулярными клетками тела мозжечка, характерен для костистых рыб, степень его развития соответствует уровню развития зрительной системы. Аналогичным образом организованы и диенцефальные проекции в тело мозжечка у телеостей, поскольку они также происходят из структур, получающих прямой сетчаточный вход. У этих-рыб. характерным является и наличие проекций в заслонку мозжечка от ядра заслонки и нижней оливы.* Нерешенным остается вопрос о существовании теленцефальных проекций в мозжечок костистых рыб, хотя отдельные сведения о них имеются.
Эфференты мозжечка лучеперых рыб, не столь многочисленные, как афференты, объединены в несколько трактов. Общими элементами структуры мозжечка для всех групп подкласса являются ножка мозжечка, brachium conjunctivum, и мозжечково-моторный тракт, tr. cerebellomotorius.
Ножка мозжечка у хондростей и голостей в основном образована аксонами нейронов его ядра и отчасти клеток Пуркинье. У телеостей эфферентными элементами являются эвридендроидные клетки ганглионарного слоя (что же касается клеток Пуркинье, то их роль в организации эфферентных церебеллофугалькых связей не доказана). Ножка мозжечка связывает его с многочисленными образованиями среднего мозга (ядрами глазодвигательного комплекса, ядром медиального продольного пучка,
торусом, тектумом, ретикулярной формацией), а также, в меньшей степени, с промежуточным мозгом. У телеостей обнаружены также входы к прекурсору красного ядра, отсутствующему у других лучеперых.
Церебелло-моторный тракт описан у представителей всех, групп лучеперых рыб. Он берет начало от тела мозжечка и (у голостей и телеостей) от заслонки. Основная часть проекций адресована продолговатому, меньшая - среднему мозгу, Значительное количество волокон проходит в составе медиального продольного пучка и достигает ядер глазодвигательного комплекса. Для некоторых групп, например хондростей, точное место окончания волокон не известно.
Некоторые группы эфферентов описаны лишь у костистых рыб. Так, для них показан мозжечково-тектальный тракт, tr. cerebellotectalis, связывающий тело мозжечка с торусом. Кроме того, у них имеются ипсилатеральные входы в октаво-латеральные центры продолговатого мозга, обеспечиваемые tr. cerebello-octavolateralis.*
Модификации строения и некоторых связей мозжечка обнаруживаются у костистых рыб отряда клюворылообразных - мормирид. У них мозжечок содержит все отделы за исключением ядра. Тело мозжечка разделено на три доли; четвертая, ray-даль'ная, составляет вестибулю-латеральную долю. Заслонка представлена складчатой структурой, покрывающей весь мозг. Основной вход в каудальную долю составлен структурами октаво-латеральной области продолговатого мозга, при этом обнаруживается пространственное разделение проекций: электросенсорные входы адресованы центральной части вестибуло-мозжечка, механорецептивные - гранулярным возвышениям. Остальные входы в мозжечок относительно немногочисленны. Спинальные афференты не обнаружены. Заслонка мозжечка получает большинство входов от нижней оливы, в которую также проецируются электросенсорные отделы продолговатого мозга. Таким образом, источник основных афферентов всех отделов мозжечка этих животных - октаво-латеральная область продолговатого мозга. Еще одной особенностью, не характерной для остальных костистых рыб, являются связи с тригеминалькым комплексом: нисходящее и среднемозговое ядра тройничного нерва формируют входы к телу мозжечка, которое в свою очередь посылает проекции к его моторному ядру.
Заключая обзор строения мозжечка рыб, следует сказать, что в пределах этого надкласса имеется немало общих черт, касающихся нейронного состава, анатомического строения, характера организации связей, что, вероятно, обусловлено общностью происхождения этого отдела и его характеристик у предковых
форм. Вместе с тем широкий спектр вариаций строения, обнаруживаемый в этом отделе, связан не только со степенью развития моторной активности, но и с общим уровнем организации нервной системы, включая особенности сенсорных входов от зрительных структур мозга - их отсутствие у хрящевых и выраженность у костистых, причем у последних также имеются вариации - хорошая выраженность входов у окунеобразных и меньшая у карпообразных. Наконец, в случае исключительно высокого развития какой-либо из сенсорных систем в строении мозжечка оказывается возможным выделить сходные черты специализации даже у представителей разных классов (например, электрические скаты и электрические костистые рыбы), что является примером параллельного и независимого преобразования мозговых отделов и иллюстрирует широкие адаптивные возможности нервной системы.
В разных группах амфибий уровень развития мозжечка значительно варьирует (см. рис. 5, 13-15). В отряде хвостатых амфибий мозжечок по строению напоминает аналогичный отдел мозга двоякодышащих: в нем различают медиальную часть и парные аурикулярные доли. Тело мозжечка также парное и соединяется слоем комиссуральных волокон. Мозжечок рудиментарен у протеевых, у других - более четко сформирован. У скрытожаберников тело мозжечка и гранулярные возвышения отделены от дна IV желудочка, и мозжечок частично скрыт производными среднего мозга. У сиреневых мозжечок представляет собой небольшое скопление клеток в передней стенке желудочка. Тело мозжечка у взрослой амбистомы трудно отграничить от гранулярного возвышения. Наконец, у созревших личинок многих тритонов мозжечок крупнее, чем у взрослой особи. Вариации строения латеральных долей связаны с образом жизни хвостатых амфибий - водным или наземным.
Нейронный состав мозжечка хвостатых амфибий весьма прост. Основные элементы представлены гранулярными клетками, имеющими типичный для всех позвоночных вид, а также примитивными клетками Пуркинье с небольшим простым дендритным полем, не имеющим преимущественной ориентации. В молекулярном слое располагаются мелкие звездчатые клетки; сведения о корзинчатых клетках отсутствуют. Эфферентными элементами являются клетки Пуркинье, а также нейроны так называемого вентрального мозжечкового возвышения, которое рассматривают как ядро мозжечка.
У бесхвостых амфибий тело мозжечка более массивно. Аурикулы, хорошо развитые у головастиков, после метаморфоза в значительной степени редуцируются и сохраняют лишь вестибулярные проекции. Во время развития бесхвостых амфибий (сем. Настоящие лягушки) мозжечок остается в незрелой стадии: наружный гранулярный слой отсутствует, слой клеток Пуркинье - тоже, хотя в пределах субвентрикулярной зоны имеются отдельные
крупные клетки. В период отрастания конечностей постепенно формируется наружный гранулярный слой путем миграции клеток от основания мозжечка, затем начинается вторая миграция, когда клетки перемещаются по направлению к желудочку. Выраженность данной миграции максимальна в период развития передних конечностей. В это же время слой клеток Пуркинье можно выделить как самостоятельный.
У взрослых животных мозжечок представлен единой гладкой пластиной, молекулярный слой которой расположен рострально и обращен к тектуму. Утраченные при метаморфозе латеральные доли превращаются в клеточную пластинку, занимающую каудальное положение. Ядро мозжечка бесхвостых амфибий достаточно четко очерчено и отделяется от комплекса вестибулярных ядер и ядра тройничного нерва проходящими трактами.
Примитивными чертами мозжечка амфибий считают аморфный слой клеток Пуркинье, а также особенности дендритного поля этих нейронов: оно не столь стереотипно, первичный ствол имеет значительную длину. Число шипиков для одной клетки составляет 3-4 тыс. (для сравнения: у обезьян - 60-120 тыс.). Лиановидные волокна подобны имеющимся у других животных: каждое волокно образует около 300 синапсов на дендрите одной клетки Пуркинье. Терминальные ветвления мшистых волокон не имеют характерных для высших позвоночных очертаний; вместо 'них отмечено рыхлое сплетение волокон, проникающих в гранулярный слой и адресованных дендритам гранулярных клеток. У амфибий, подобно другим животным, аксоны гранулярных клеток формируют в молекулярном слое систему параллельных волокон. В молекулярном слое присутствуют и звездчатые клетки, на теле и дендритах которых имеются синаптические контакты.*
"Мшистые волокна" амфибий образованы аксонами нейронов экстрацеребеллярных структур. К ним относятся: вестибулярный комплекс, ядро базального оптического тракта, дорсальные рога спинного мозга. Небольшая часть представлена волокнами дорсальных канатиков. В ранних работах описаны тригемино- и лобо-церебеллярный входы, последний характерен для личиночной стадии. Тройничный вход экспериментально не подтвержден. Имеются сведения и о проекции в мозжечок моторных ядер глазодвигательного комплекса, языкоглоточного и лицевого нервов.
Лиановидные волокна, исходя из сходства в характере образуемых ими контактов с имеющимися у высших позвоночных, рассматривают как афференты от нижней оливы. Кроме того, в
области аурикул они являются волокнами VIII нерва. Эфференты у бесхвостых амфибий представлены аксонами нейронов единственного ядра. Волокна после перекреста в пределах мозжечка образуют его ножку. Основной объем проекций мозжечка адресуется нижележащим структурам: ретикулярной формации среднего и продолговатого мозга, в том числе области при-мордиального красного ядра, и вестибулярному комплексу, а также, возможно, спинному мозгу. Восходящие в таламус проекции недавно показаны у Rana.
Таким образом, у амфибий мозжечок содержит клеточные элементы, свойственные этому отделу мозга высших позвоночных, с аналогичным характером взаимодействий между ними: возбуждающее влияние оказывают мшистые, лиановидные, параллельные волокна; тормозное - клетки Гольджи, звездчатые клетки, коллатерали аксонов клеток Пуркинье. Вместе с тем, у амфибий еще отсутствуют характерные для высших позвоночных гломерулы и корзинчатые сплетения, невелико число параллельных волокон, дендриты клеток Пуркинье бедны шипиками, и ориентация их ветвлений не столь закономерна. Особенностью связей мозжечка некоторых бесхвостых амфибий являются входы в таламус, хотя объем их невелик.
* Доказательства их гомологичное известным спинно-мозжечковым трактам млекопитающих отсутствуют.
** Отметим, что до сих пор весьма малочисленны данные об источниках и областях окончания мезенцефало- и лобо-церебеллярных трактов. Предполагавшийся вход от области представительства органов боковой линии в тело мозжечка экспериментально не подтвержден.
* Некоторые авторы отрицают ее наличие в мозжечке многоперов, другие же считают крупный, лежащий субтектально отдел мозжечка именно заслонкой, а не телом.
* В заслонку мозжечка поступают также проекции ядра Ш нерва, переднего нерва боковой линии и прекурсора красного ядра. Наличие входов от октаво-латеральной системы расценивалось некоторыми авторами как свидетельство гомологичности заслонки костистых рыб аурикулам пластиножаберных. Другие исследователи рассматривали заслонку как ростральную часть тела мозжечка и использовали ее как пример "церебелляризации" мозга рыб. Наконец, существует мнение о независимости происхождения этого отдела мозжечка, не имеющего гомологов у других позвоночных.
* Для карпа обнаружены также связи заслонки с гипоталамусом и долей лицевого нерва.
* Система параллельных волокон в мозжечке амфибий не столь объемна: лишь 3-5 тыс. волокон пронизывают дендритное поле одной клетки Пуркинье. Более крупные звездчатые клетки еще не имеют типичных "корзинок", хотя их аксоны проходят вдоль дольки в молекулярном слое.
§ 2. МОЗЖЕЧОК ВЫСШИХ ПОЗВОНОЧНЫХ
Прикрепленный к стволу мозга мозжечок рептилий имеет две пары ножек - передние и задние, pedunculi anterior et posterior. Его тело состоит из двух клеточных масс, соединенных срединной частью. Следы передней и задней борозд, подобных присутствующим в раннем эмбриогенезе у птиц и млекопитающих, обнаружены у всех рептилий и наиболее выражены в мозге хамелеонов, игуан и особенно крокодилов во взрослом состоянии. Этими бороздами мозжечок делится на передний, средний и задний отделы.
Мозжечок рептилий разнообразен, как по размерам, так и по форме. У черепах он закрывает ростральную часть IV желудочка. У многих ящериц, особенно у варанов и хамелеонов, он смещается вперед, закрывая каудальный отдел среднего мозга, причем эвертированный характер его приводит к тому, что гранулярный слой оказывается на поверхности мозжечка. У многих ящериц, тем не менее, мозжечок мал, а у амфисбен состоит из тонкой пластинки, покрывающей IV желудочек. Простым строением характеризуется и мозжечок змей (см. рис. 6).
"Аурикулярные доли" рептилий развиты незначительно. В наиболее выраженной форме они имеются у крокодилов и состоят из небольших латеральных скоплений нервной ткани, расположенных над латеральным углублением IV желудочка, и тонкой пластинки, идущей по заднему краю тела мозжечка. Последнюю рассматривают как гомолог узелковой дольки мозжечка
млекопитающих, а латеральные отделы гомологизируют с клочком. У многих других рептилий (змеи, ящерицы, черепахи) эта часть мозжечка представлена латеральными участками гранулярного слоя.*
В мозжечке рептилий выделяют медиальное и латеральное ядра, nucll. cerebelli medialis et lateralis. Медиальное располагается в базальной части тела мозжечка, дорсально от IV желудочка. Латеральное ядро у некоторых видов не отделено or медиального. Рострально ядра мозжечка граничат с ядром перешейка.
Гистологическая структура мозжечка рептилий типична для: всех позвоночных; выделяют молекулярный, ганглионарный и гранулярный слои. Мшистые волокна, приходящие из субцеребеллярных отделов, образуют контакты с гранулярными клетками. Лиановидные волокна, повторяя ход ветвлений дендритов клеток Пуркинье, заканчиваются на их шипиках; предполагается их оливарное происхождение (по аналогии с доказанным у птиц и млекопитающих). Гранулярные клетки посылают аксоны в молекулярный слой, где формируются типичные параллельные волокна, пронизывающие под прямым углом плоскость ветвления дендритов клеток Пуркинье. Параллельные волокна, так же как мшистые и лиановидные, формируют возбуждающие входы на клетки Пуркинье.
В мозжечке рептилий обнаруживаются и интернейроны, характерные для мозжечка птиц и млекопитающих. В молекулярном слое располагаются звездчатые и примитивные корзинчатые клетки (описаны у хамелеонов и ящериц). В гранулярном слое присутствуют клетки Гольджи. Помимо тормозных входов которые образуют аксоны этих нейронов, в мозжечке рептилий имеются тормозные катехоламинергические волокна.
Для всех млекопитающих, птиц и некоторых рептилий характерно наличие так называемой коры мозжечка, образующейся в результате миграции клеток к поверхности, в отличие от низших позвоночных, где клетки распределены по всей толще стенок мозжечка. Кора мозжечка отделена от перивентрикулярной области скоплением проводящих путей, составляющим белое: вещество мозжечка. Весь мозжечок птиц, млекопитающих и, среди рептилий, крокодилов поделен многочисленными бороздами на десять долек, объединенных в три доли: переднюю (I-V), среднюю (VI-VIII) и заднюю (IX и X). Дольки варьируют как по размерам, так и по внутренней структуре. Так, у некоторых птиц (например орел, пингвин) в области I и X долек практически отсутствует белое вещество, и гранулярные клетки: отделены от вентрикулярной поверхности лишь эпендимным слоем.
В сравнении с другими отделами очень мал молекулярный слой X дольки (клочок, или флоккулус). У всех птиц невелика долька III, у пингвинов мала долька I, а у многих млекопитающих обнаруживается редукция дольки I, которая еще велика у утконоса. У млекопитающих, относящихся к плацентарным, происходит значительное увеличение латеральных отделов дольки V.
У плацентарных мозжечок связан со стволом тремя парами кожек. Нижние ножки мозжечка, pedunculi cerebellares inferiores, содержат в основном афферентные волокна и формируются с уровня продолговатого мозга, в пределах которого носят название веревчатых тел, corporae restiformes. Средние ножки, pedunculi cerebellares medii, - специфическая структура мозжечка млекопитающих - связывают мозжечок с мостовыми ядрами. Верхние ножки, pedunculi cerebellares superiores, в основном составлены волокнами соединительной ножки мозжечка, имеющейся у всех позвоночных, и содержат преимущественно-мозжечковые эфференты. Процессы специализации у разных млекопитающих отражаются и на внешнем строении мозжечка. Так, у китов чрезвычайно развиты дольки IX и X, у свиней при незначительной выраженности дольки I хорошо представлена долька II, у хищных и приматов разрастаются латеральные отделы долек V и VI.
Другой особенностью высших позвоночных является хорошее развитие ядер мозжечка. У большинства птиц они представлены двумя крупными скоплениями клеток - медиальным и латеральным ядрами. Первое гомологично одноименному ядру рептилий, а значительная часть латерального - латеральному ядру высших рептилий. Однако у многих видов птиц помимо этих ядер выделяются и группы диффузно расположенных клеток. Этим обстоятельством отчасти объясняются имеющиеся расхождения в классификации ядер и их гомологизации с ядрами мозжечка млекопитающих.
У млекопитающих в медиолатеральном направлении в пределах мозжечка выделяют четыре ядра: медиальное, или ядро шатра, nucl. medialis cerebelli (nucl. fastigii), переднее и заднее промежуточные ядра, nucll. intermedius anterior et posterior (у человека - пробковидное и шаровидное соответственно, nucll. emboliformis et globosus), и латеральное, nucl. lateralis (у высших - зубчатое, nucl. dentatus). Медиальное ядро гомологично одноименному ядру мозжечка птиц, промежуточное - дорсальной части, а латеральное - вентролатеральной части латерального ядра птиц. Латеральное ядро мозжечка достигает у приматов значительных размеров и имеет сложную форму, в то время как другие ядра относительно меньше.
Строение мозжечка у всех высших позвоночных удивительно сходно. Каждая его долька во внутренних отделах представлена белым веществом, а снаружи покрыта трехслойной корой,
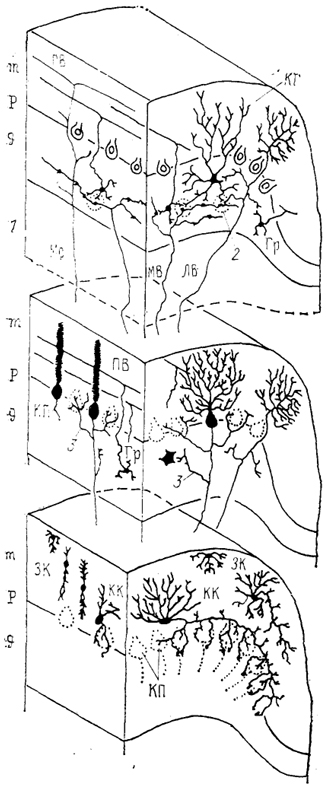
Рис. 46. Строение коры мозжечка приматов (по: Haines, 1986).
1 - белое вещество, 2 - гломерулы, 3 - коллатерали аксонов клеток Пуркинье. ЛВ, MB, Mс, ПВ - лиановидные (ЛВ), мшистые (MB), мультислойные (Мс), параллельные (ПВ) волокна. Остальные обозначения, как на рис. 45.
При этом у некоторых рептилий ганглионарный слой составлен из нескольких рядов клеток Пуркинье, у других, подобно птицам и млекопитающим,- только одним рядом грушевидных клеток (рис. 46).*
Афферентами коры мозжечка служат мшистые и лиановидные волокна. Первые составлены в основном аксонами нейронов прецеребеллярных отделов, исключение составляет лишь их небольшая часть - входы мозжечковых ядер. Источником формирования лиановидных волокон являются ядра нижней оливы.
Гранулярный слой, str. granulare, - самый внутренний слой в коре (кроме эвертированного мозжечка у рептилий), в основном составлен плотно лежащими телами мелких округлых клеток-зерен, а также клетками Гольджи. Здесь же проходят мшистые и лиановидные волокна (рис. 47). Гранулярные клетки наиболее многочисленны (у человека - 1010-1011). Размеры клеток-зерен невелики: у высших млекопитающих их диаметр 5-7 мкм, у низших млекопитающих и рептилий несколько больше - 7-9 мкм. Их редкие короткие дендриты ветвятся вблизи сомы, формируя когтевидные окончания. Аксоны начинаются от тела или от проксимальных участков одного из дендритов, восходят в молекулярный слой и там, Т-образно делясь,
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|