- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 1 страница
Эволюционная морфология нервной системы позвоночных: Учебник для студентов вузов. Андреева Н.Г., Обухов Д.К. Изд. 2-е, доп., изм. М.: Лань, 1999. 384c. ISBN 5-8114-0074-8

ОГЛАВЛЕНИЕ
| Предисловие | |
| Список условных сокращений | |
| Глава 1. Общая характеристика нервной системы позвоночных (Д.К. Обухов) | |
| § 1. Общие черты строения и эмбрионального развития нервной системы позвоночных | |
| § 2. Система желудочков мозга | |
| § 3. Уровни и принципы организации нервной системы | |
| Глава 2. Спинной мозг (Д.К. Обухов) | |
| § 1. Общие представления об организации спинного мозга позвоночных | |
| § 2. Проводящие пути спинного мозга | |
| Глава 3. Продолговатый мозг (Н.Г. Андреева) | |
| § 1. Общий план строения продолговатого мозга | |
| § 2. Сенсорные ядра продолговатого мозга | |
| § 3. Моторные ядра продолговатого мозга | |
| § 4. Ядра области покрышки продолговатого мозга | |
| Глава 4. Мозжечок (Н.Г. Андреева) | |
| § 1. Мозжечок низших позвоночных | |
| § 2. Мозжечок высших позвоночных | |
| Глава 5. Средний мозг (Н.Г. Андреева) | |
| § 1. Сенсорные ядра среднего мозга | |
| § 2. Моторные ядра среднего мозга | |
| § 3. Ядра области покрышки среднего мозга | |
| Глава 6. Ретикулярная формация (Н.Г. Андреева) | |
| § 1. Медианная зона ретикулярной формации | |
| § 2. Медиальная зона ретикулярной формации | |
| § 3. Латеральная зона ретикулярной формации | |
| § 4. Сравнительный анализ ретикулярной формации позвоночных | |
| Глава 7. Промежуточный мозг (Н.Г. Андреева) | |
| § 1. Структурная организация таламуса | |
| § 2. Общая характеристика гипоталамуса | |
| § 3. Эпиталамус | |
| Глава 8. Конечный мозг (Д.К. Обухов) | |
| § 1. Конечный мозг низших позвоночных | |
| § 2. Конечный мозг высших позвоночных | |
| Глава 9. Функциональные аспекты преобразования нервной системы (Н. Г. Андреева) | |
| Рекомендуемая литература | |
ПРЕДИСЛОВИЕ
Последние два десятилетия можно считать периодом определенного подъема сравнительной нейроморфологии. Это связано, во-первых, с появлением принципиально новых морфологических и экспериментально-морфологических методик исследования нервной ткани и межнейронных связей, позволивших получить ряд фактов, которые изменили наши представления об организации центральной нервной системы позвоночных и путях ее эволюционных преобразований. Во-вторых, расширение видового состава изучаемых представителей современных позвоночных показало, что имеется множество вариантов строения нервных центров, причем даже у близкородственных видов различия могут оказаться весьма значительными, при относительном сходстве функциональных возможностей животных.
Это наряду с данными сравнительной морфологии, палеонтологии, систематики и других дисциплин свидетельствует о неравномерном, во многом противоречивом характере эволюции ЦНС, подчеркивает необходимость дифференцированного подхода к интерпретации данных, получаемых на представителях разных групп позвоночных.
Необходимо отказаться от традиционных линейных схем эволюции нервной системы по типу "от низших к высшим". В каждой группе современных позвоночных имеются различные варианты организации нервных центров и мозга в целом, развивающиеся параллельно и независимо друг от друга. В этом плане даже мозг человека представляет собой один из возможных и реализуемых в эволюции вариантов организации ЦНС.
Большая часть руководств, монографий, обзорных статей, посвященных данной теме, труднодоступна студентам, а соответствующей учебной литературы нет. Таким образом возникла необходимость создания учебного пособия, в котором систематизировались бы данные по морфологии мозга позвоночных, полученные за последние годы, и основное внимание уделялось
бы представителям домлекопитающих животных, которые все чаще становятся объектами лабораторных исследований.
Пособие написано на основе курсов лекций по морфологии и эволюции нервной системы позвоночных, читаемых для студентов кафедр физиологии высшей нервной деятельности и цитологии и гистологии СПбГУ. Необходимо отметить, что пособие рассчитано на подготовленного читателя, знакомого с вопросами общей организации нервной системы, сравнительной анатомии и физиологии, палеонтологии и систематики позвоночных.
Ограниченный объем книги и неограниченный объем информации не дали возможности подробно разобрать (а в некоторых случаях даже упомянуть) ряд вопросов организации нервной системы, в частности вегетативной нервной системы, нейро-эндокринной системы, органов чувств. Частично это может быть компенсировано наличием по этим проблемам современной учебной литературы.
Данное пособие предназначено студентам биологических и физиологических кафедр университетов, медицинских и педагогических институтов, а также может быть полезно аспирантам и молодым нейроморфологам и нейрофизиологам, интересующимся вопросами организации и эволюции нервной системы. Авторы надеются, что оно будет способствовать формированию правильного биологического подхода к решению сложных вопросов организации и эволюции ЦНС.
Авторы будут признательны за критические замечания и пожелания.
СПИСОК УСЛОВНЫХ СОКРАЩЕНИЙ
- А - amygdala
- АР- area preoptica
- АРН - area parahippocampalis
- aq - aqueductus cerebri
- Ast - archistriatum
- AT - area triangularis thalami
- aur - a uricula cerebelli
- AVT a rea ventralis tegmenti
- BO - bulbus olfactorius
- ca - commissura anterior
- Caj - nucleus (nucl.) interstitialis Cajal
- cc - corpus callosum
- cCer - corpus cerebelli
- Cd - nucl. caudatus
- CDL - area corticoidea dorsolateralis
- Cer - Cerebellum
- Ceri - nucl. interpositus cerebeili
- Cerl - ,, lateralis ,,
- Germ - ,, medialis ,,
- CGL - corpus geniculatum late-rale
- CGM - ,, ,, mediale
- ch - chiasma
- cha - commissura habenularis
- Co - nucl. cochlearis
- Coa - ,, ,, angularis
- Colm - ,, ,, laminaris
- Com - ,, ,, magnocellularis
- CP - ,, centralis posterior thalami
- cp - commissura posterior
- crCer - crista cerebelli
- Crx - cortex cerebri
- Cs - colliculus superior
- ct - commissura tecti
- Cun - nucl. cuneatus
- Cuna - ,, ,, accessorius
- Dien - Diencephalon
- DL - nucl. dorsalis lateralis thalami
- DM - ,, ,, medialis ,,
- DO - ,, octavus descendens
- Dol - ,, dorsalis areae octavo-lateralis
- DP - ,, ,, posterior thalami
- E - Epiphysis
- EM - nucl. ectomammillaris
- Ep - ,, entopeduncularis
- Est - ectostriatum
- Eth - eminentia thalami
- Fl - flocculus cerebelli
- Flm - nucl. fasciculi longitudi-nalis medialis
- flm - fasciculus longitudinalis
- medialis
- FR - formatio reticularis
- FR1 - ,, ,, pars lateralis
- FRm - ,, ,, ,, medialis
- ftl - fasciculus telencephalicus lateralis
- ftm - ,, ,, medialis
- G - nucl. gracilis
- g - stratum granulare cere-belli
- Gl - nucl. glomerulosus
- GP - globus pallidus
- GS - nucl. gustatorius secundarius
- gs - tractus (tr.) ,, ,,
- HA - hyperstriatum accessorius
- Ha - habenula
- HD - hyperstriatum dorsale
- HDM - nucl. dorsomedialis hypothalami
- HIS - hyperstriatum intercalates accessorius
- HL - hypothalamus area lateralis
- HM - ,, ,, medialis
- HP - ,, ,, posterior
- HPv - ,, ,, periventricularis
- HV - hyperstriatum ventrale
- HVM - nucl. ventromedialis hypothalami
- Ну - hypothalamus
- Hyd - ,, pars dorsalis
- hyp - hypophysis
- Hyv - hypothalamus pars ven-tralis
- I - nucl. isthmi
- ICo - ,, intercollicularis
- Im - ,, isthmi magnocellularis
- Int - ,, intermedius areae octavolateralis
- IP - ,, interpeduncularis
- Ip - ,, isthmi .parvocellularis
- LC - locus coeruleus
- LI - lobus (lob.) inferior hypothalami
- Ll - nucl. lemnisci lateralis
- ll - lemniscus lateralis
- Lll - lob. lineae lateralis
- Llla - ,, ,, ,, anterior
- Lllp - ,, ,, ,, posterior
- 1m - lemniscus medialis
- LPO - lob. Parolfactorius
- Lsp - ,, spinalis
- LVII - ,, facialis
- LIX - ,, glossopharyngei
- LX - ,, vagi
- m - stratum moleculare cerebelli
- MD - nucl. dorsomedialis thalami
- Mes - Mesencephalon
- Met - Metencephalon
- Mo - Medulla oblongata
- Mol - nucl. octavolateralis medialis
- Msp - Medulla spinalis
- nA - nucl. anterior thalami
- na - ,, accumbens
- nB - ,, Bellonchi
- nC - ,, centralis thalami
- nCer - ,, cerebelli
- Ncx - neocortex
- nDI - nucl. dorsolateralis areae octavolateralis
- nL - ,, lateralis thalami
- nlla - nervus anterior lineae lateralis
- nllp - ,, posterior ,,
- noa - ,, olfactorius anterior
- nP - ,, posterior thalami
- nR - ,, ruber
- nSp - nervi spinales
- Nst - neostriatum
- nTs - nucl. tractus solitarii
- nV - ,, ventralis thalami
- n III - n XII nervi craniales
- Oi - oliva inferior
- Os - ,, superior
- P - stratum ganglionare cerebelli
- PA - paleostriatum augmentatum
- Pav - nucl. paraventricularis
- Pb - ,, parabrachialis
- PD - pallium dorsale
- pGl - nucl. preglomerulosus
- PL - pallium laterale
- PM - ,, mediale
- Pn - pons
- PO - nucl. preopticus
- PP - paleostriatum primitivum
- Prof - nucl. profundus mesencephali
- Pst - paleostriatum
- Pt - nucl. pretectalis
- Pul - pulvinar
- Put - putamen
- r - nucl. raphes
- Ret - ,, reticularis thalami
- Reu - ,, reunions ,,
- Rh - Rhombencephalon
- Ri - nucl. reticularis inferior
- ri - ,, raphes ,,
- RL - ,, reticularis lateralis
- rl - ,, raphes linearis
- Rm - ,, reticularis medius
- rm - ,, raphes magnus
- ro - ,, ,, obscurus
- Rot - ,, rotundus
- rp - ,, raphes pallidus
- rs - ,, ,, superior
- Rs - ,, reticularis superior
- Rsl - ,, ,, ,, lateralis
- Rsm - ,, ,, ,, medialis
- scd - tr. spinocerebellaris dorsalis
- Sch - nucl. suprachiasmaticus
- scv - tr. spinocerebellaris ventralis
- Sg - nucl. suprageniculatus
- Sgc - substantia grisea centralis mesencephali
- Si - ,, innominata
- sm - stria medullaris
- Sn - substantia nigra
- Sp - nucl. septalis
- Spl - ,, ,, lateralis
- Spm - ,, ,, medialis
- Srh - sulcus rhynalis
- sRot - nucl. subrotundus
- Str - striatum
- Strd - ,, dorsale
- Strv - ,, ventrale
- Sv - Saccus vasculosus
- Syn - synencephalon
- tb - tr. tectobulbaris
- tbd - ,, ,, dorsalis
- Tel - Telencephalon
- Tg - tegmentum mesencephali
- Tgl - nucl. tegmenti lateralis
- Tgv - ,, ,, ventralis
- Th - thalamus
- ThD - ,, pars dorsalis
- Thd - ,, dorsalis
- Thdl - nucl. dorsolateralis thalami
- Thdm - dorsomedialis
- ThV -thalamus pars ventralis
- Thv - ,, ventralis
- Thvl - nucl. ventrolateralis thalami
- Thvm - ,, ventromedialis ,,
- Tl - torus longitudinalis
- TO -tectum opticum
- Tol -tuberculum olfactorium
- TP - ,, posterior
- TPC - nucl. tegmenti pedunculopontinus pars compacta
- trs - tr. rubrospinalis
- Ts - torus semicircularis
- Tsc - ,, ,, , nucl. centralis
- Tsl - ,, ,, , ,, laminaris
- Va - valvula cerebelli
- Vedl - nucl. vestibularis dorsolateralis
- Veds - ,, ,, descendens
- Vel - ,, ,, lateralis
- Vem - ,, ,, magnocellularis
- Ves - ,, ,, superior
- Vest - nucl. vestibulares
- Vetg - nucl. vestibularis tangentialis
- Vevl - ,, ,, ventrolateralis
- Vevm - ,, ,, ventromedialis
- VL - ventrolateralis thalami
- VM - nucl. ventromedialis thalami
- vO - ventriculus opticus
- Vol - nucl. ventralis areae octavolateralis
- vsl - tr. vestibulospinalis
- III - nucl. motorius nervi oculomotorii
- IIIa - ,, accessorius ,, ,,
- IV - ,, motorius nervi trochlearis
- V - ,, descendens (spinalis) nervi trigemini
- Vm - ,, motorius ,, ,,
- Vmes - mesencephalicus ,, ,,
- Vpr - ,, sensorius principalis
- VI - ,, motorius nervi abducentis
- VIa - ,, accessories ,, ,,
- VIIm - ,, motorius nervi facialis
- VIIIa - ,, octavus anterior
- VIIIm - ,, ,, medius
- VIIIp - ,, ,, posterior
- IXm - ,, motorius nervi glossopharyngei
- X - ,, nervi vagi
- Xdm - ,, dorsalis motorius nervi vagi
- Xm - ,, motorius ,, ,,
- XI - ,, nervi accessorii
- XII - ,, nervi hypoglossi
ГЛАВА 1
ОБЩАЯ ХАРАКТЕРИСТИКА НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ
Становление типа хордовых, Chordata, шло в докембрии, по-видимому, в период, когда примитивные оболочники (подтип Tunicata), обладающие личинкой, переходили к неотении и потере прикрепленной стадии. Бесчерепные (подтип Acrania), произошедшие от таких форм, в ходе морфофизиологического развития дали начало позвоночным (подтип Vertebrata) (рис. 1).
Современные позвоночные представляют собой конечные этапы широкой и многоступенчатой адаптивной радиации. Они довольно рано начали утверждаться на высших ступенях трофических пирамид, что давало каждой отдельной группе определенные преимущества, но при этом обусловливало и ряд сложностей, например в добывании пищи. Экологическая дифференциация позвоночных, сопровождающаяся у разных форм интенсификацией функций органов, определялась прогрессивным усложнением среды их обитания. Таким образом усилился отбор по способ'ности адекватно отвечать на весьма разнообразные изменения среды, что обеспечивалось прежде всего развитием нервной системы, поскольку частные приспособления в отличие от преобразований ЦНС не имеют столь универсального значения в изменчивой и сложной обстановке. При этом процессы усложнения и дифференциации нервной системы и ее структурных признаков, протекая в сходных условиях, в филогенетически далеких ветвях позвоночных могли приводить к формированию в ЦНС функционально сходных, но отличающихся по своему происхождению структур (конвергенция). Процессы же усложнения среды обитания при наличии общей наследственной основы у близкородственных форм могли в еще большей степени определить-однонаправленные преобразования в ЦНС. Параллельные преобразования в нервной системе распространены намного шире, хотя в близкородственных группах трудно выявить результаты таких параллельных, независимых изменений.
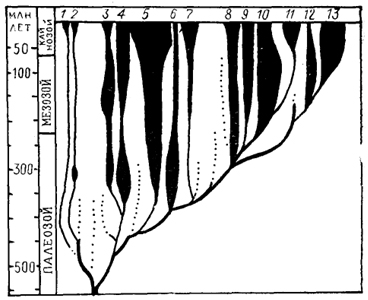
Рис. 1. Схема эволюции основных классов позволенных (по: Fite, 1985).
1 - бесчерепные; 2 - круглоротые; 3-7 - рыбы: цельноголовые (3), пластиножаберные (4), лучеперые и многоперые (5), саркоптеригии (6), двоякодышащие (7); 8 - амфибии; 9-11 - рептилии: черепахи (9), чешуйчатые (10), архозавры (11); 12, 13 - млекопитающие: первозвери (однопроходные) (12), звери (сумчатые и плацентарные) (13).
До недавнего времени характеристику отдельных групп позвоночных давали на примере небольшого числа их типичных представителей - "модельных" форм. Вместе с тем, "модельные" формы не всегда отражают все многообразие признаков, конкретного уровня организации. Данные палеонтологии и сравнительной анатомии показывают, что наиболее перспективным при изучении эволюции той или иной группы позвоночных является, с одной стороны, исследование так называемых экологически консервативных реликтов, обитающих в среде, по абиотическим и биотическим характеристикам мало отличающейся от среды обитания их предков, а с другой стороны, исследование возможно большего количества современных видов. При этом необходимо учитывать вероятность значительной неравномерности хода эволюции и определенную "мозаичность" преобразования разных структур и признаков (в том числе и ЦНС) в линии предок - потомок. Формирование признаков проходило неравномерно и нередко параллельно, независимо, даже в близкородственных группах. При этом "принцип мозаичности" может проявляться как в сохранении" у одних представителей группы тех или иных черт древнего, "исходного", состояния, так и в возникновении у других широкого спектра признаков иных уровней организации, часто более высоких, чем у "модельных" форм. Это явление в эволюции позвоночных, так называемое " филогенетическое опережение", есть реальный результат неравномерности эволюционного процесса, характерной для всех уровней организации живого.
§ 1.ОБЩИЕ ЧЕРТЫ СТРОЕНИЯ И ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
НЕРВНОЙ СИСТЕМЫ ПОЗВОНОЧНЫХ
Нервная система всех позвоночных развивается из элементов наружного зародышевого листка-эктодермы. Этот процесс имеет определенные особенности у представителей разных
групп, однако ему свойственны и общие для всех позвоночных закономерности.
Начальный этап развития нервной системы состоит в том, что на дорсальной стороне зародыша обособляется участок эктодермы - нервная пластинка, элементы которой интенсивно размножаются и дифференцируются, превращаясь в узкие цилиндрические нейроэпителиалькые клетки, отличные от соседних клеток покровного эпителия. В результате интенсивного деления и неравномерного роста нейроэпителия происходит его инвагинация с последующим формированием нервной трубки. Замыкание нервной трубки начинается в середине зародыша, затем процесс распространяется к головному и хвостовому концам эмбриона, где некоторое время остаются незамкнутыми отверстия - передний и задний нейропоры.
Основной причиной формирования нервной пластинки и замыкания ее s нервную трубку является преобразование нейроэпителиальных клеток, связанное с изменением ориентации компонентов их цитоскелета. Нервная трубка (как полагают, под индуцирующим воздействием хорды) постепенно погружается в мезодерму зародыша и под влиянием мезодермальных сомитов разделяется на сегментарные участки - нейромеры. Сомиты располагаются по сторонам нервной пластинки и вдавливаются в нее, определяя конфигурацию будущих нейромеров (рис. 2).
У всех позвоночных развитие первой пары головных сомитов связано с формированием переднего мозга, но дальнейший процесс сегментации нервной системы идет по-разному у представителей разных групп позвоночных. Так, у хрящевых рыб и амфибий каждый последующий головной сомит соответствует по объему двум сомитам костистых рыб и амниот.
В дальнейшем головные сомиты сливаются и образуют три основных -сегмента: премандибулярный, мандибулярный и гиоидный. Границей головных сегментов служит область ушной капсулы, за которой формируются от 2-3 до 10-12 туловищных сегментов (в зависимости от группы позвоночных). Параллельно формируется система черепномозговых нервов. Каждый сегмент иннервируется определенными парами нервов: премандибулярный - nervus terminalis и глазодвигательным нервом (III); мандибулярный-тройничным (V) и блоковым (IV) нервами; гиоидный - отводящим (VI) и лицевым (VII) нервами. Следующие за головными два сегмента иннервируются соответственно языкоглоточным (IX) и блуждающим (X) нервами. Ростральные туловищные сомиты у высших позвоночных иннервируются системой .добавочного нерва (XI), включающего в себя разное количество корешков в зависимости от числа туловищных сомитов. Подъязычный нерв (XII), ин-нервирующий гипобранхиальную мускулатуру, которая развивается из закладки туловищных сегментов, по своей функции аналогичен вентральным (соматомоторным) корешкам спинномозговых нервов, иннервирующих поперечнополосатую мускулатуру туловища и конечностей.
Еще на стадии замыкания нейропоров начинается ростро-каудальная дифференцировка нервной трубки зародыша. Передний конец трубки значительно расширяется, боковые стенки "утолщаются, образуя зачатки мозговых пузырей. Лежащий краниально пузырь образует первичный передний мозг, Prosencephalon,
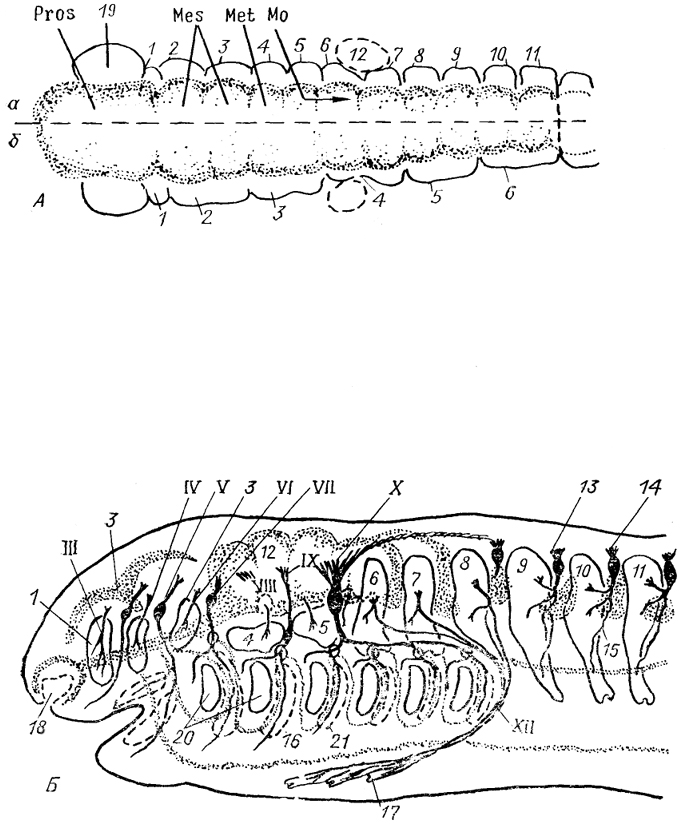
Рис. 2. Схема развития мозговых пузырей (A) и формирования головной части зародыша позвоночных (Б)(по: Northcutt, 1979; Jacobson, 1988).
А: а, б - сомитомеры и мозговые пузыри костистых рыб и амниот (а), хрящевых рыб и амфибий (б). 1-11 - сомитомеры (будущие туловищные сомиты); 12 - закладка ушной капсулы; 13 - спинальный ганглий; 14, 15 - дорсальный (14) и вентральный (15) корешки спинномозгового нерва; 16 - полость целома; 17- гипобранхиальная мускулатура; 18 - обонятельный орган, 19- глазной пузырь; 20, 21 - жаберные щели (20) и дуги (21); III-X, XII - черепномозговые нервы. Здесь и далее остальные обозначения см. в списке условных сокращений.
средний пузырь - первичный средний мозг, Mesencephalon, а из третьего пузыря, который переходит в закладку спинного мозга, развивается первичный задний мозг, Rhombencephalon. Вскоре первый и третий мозговые пузыри с помощью
борозд-сужений разделяются, образуя каждый по два вторичных мозговых пузыря. Эта стадия развития головного мозга получила название стадии пяти мозговых пузырей. Самым ростральным отделом нервной трубки становится конечный мозг, Telencephalon. Каудальнее располагается промежуточный мозг, Diencephalon, за которым следует средний мозг, Mesencephalon. Первичный задний мозг разделяется на вторичный задний мозг, Metencephalon, и продолговатый, Medulla oblongata.
Спинной мозг, Medulla spinalis, образуется из каудальных: отделов нервной трубки. Он представляет собой часть ЦНС, в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных: трубчатый характер строения и сегментарность.
После формирования мозговых пузырей в нервной системе начинаются сложные процессы внутренней дифференцировки и роста. Уже на ранних этапах развития зародыша нервная трубка на значительном протяжении разделяется проходящей по вентрикулярной поверхности пограничной бороздой, sulcus limitans, на два отдела: дорсальный - крыловидная пластинка, и вентральный - базальная пластинка. Участки мозга, развивающиеся из крыловидной пластинки, содержат сенсорные ядра, из базальной - моторные и вегетативные; ростральная часть не содержит базальной пластинки и целиком происходит из крыловидной. Отделы головного мозга, содержащие-производные обеих пластинок - средний, задний, продолговатый- часто объединяют названием "ствол мозга".
Разделение головного мозга на пять отделов (происходящих из мозговых пузырей) характерно для представителей всех групп позвоночных - от круглоротых до млекопитающих и человека, хотя структура и функциональная организация этих отделов претерпевают в эволюции позвоночных значительные изменения (рис. 3).*
Продолговатый мозг, Medulla oblongata,является наиболее каудальным отделом головного мозга, плавно переходящим в спинной. Условной границей головного и спинного мозга считают место выхода первой пары спинномозговых нервов. Наиболее значительное сходство со спинным мозгом наблюдается в каудальных отделах продолговатого мозга и выражается в расположении серого и белого вещества, а также в центральном положении мозгового канала. Последний в ростральном направлении расширяется и превращается в ромбовидную или треугольную полость - IV мозговой желудочек, ventriculus-quartus. Крыша его истончена и составлена лишь эпендимным эпителием, снаружи которого у высших позвоночных лежит
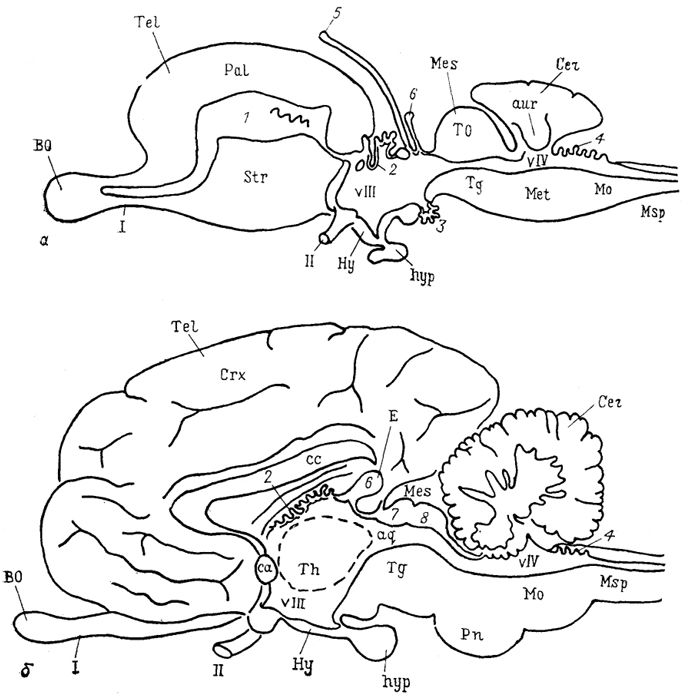
Рис. 3. Схема общей организации головного мозга низших (а) и высших (б) позвоноччых (по: Plate!, 1984).
1-4 - сосудистые сплетения конечного (1), промежуточного (2, 3),заднего (4) мозга; 5- теменной орган; 6 - пинеальный орган (эпифиз); 7, 8 - верхние (7) и нижние (8) бугорки четверохолмия среднего мозга; vIII, vIV - желудочки промежуточного (vIII) в заднего (vIV) мозга; I, II - обонятельный (I) и зрительный (II) черепномозговые нервы.
сильно васкуляризованная мягкая оболочка мозга. У низших позвоночных серое вещество представляет собой тяжи, или столбы, нервных клеток, идущие вдоль продолговатого мозга, тогда как у высших позвоночных намечается четкая тенденция к разделению этих тяжей на отдельные ядра.
Задний мозг, Metencephalon,у всех позвоночных представлен мозжечком, Cerebellum, форма, размеры и соотеошение частей которого сильно варьируют у представителей разных
групп позвоночных и в целом коррелируют со степенью совершенства его моторных реакций.
В вентральной и боковых стенках заднего мозга, слабо развитых у низших позвоночных, у млекопитающих формируется: мощная система проводящих трактов и ядер, образующая самостоятельный отдел -: мост, Pons. Полостью заднего мозга; являются ростральные отделы IV мозгового желудочка.
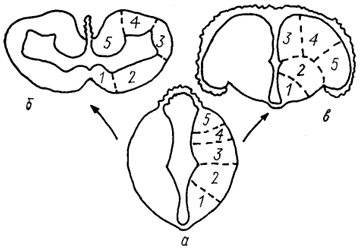
Рис. 4. Два типа развития конечного мозга позвоночных.
а - ранняя стадия развития; б, в - инвертированный (б)и эвертированный (а) типы строения мозга. 1 - стриатум; 2 - септум; 3-5 - латеральная (3), дорсальная (4) и медиальная (5) зоны паллиума.
Средний мозг, Mesencephalon, у низших позвоночных в дорсальных отделах сформирован крупными зрительными долями (tectum opticum), полости которых - зрительные желудочки соединяются с общей полостью среднего мозга - водопроводом мозга, aqueductus-cerebri. У млекопитающих на месте зрительных долей: развиваются структуры четверохолмия, под которым; располагается зона покрышки. Вентральную часть среднего мозга у высших позвоночных занимают ножки Мозга - система трактов, связывающих вышележащие отделы ЦНС с нижележащими.
Промежуточный мозг, Diencephalon, включает в себя лежащий дорсально эпиталамус, epithalamus, срединную часть - таламус, thalamus, и расположенный вентрально гипоталамус, hypothalamus. У высших выделяют несколько образований, формирующих субталамус, subthalamus, генез которого до конца неясен.
Конечный мозг, Telencephalon, представляет собой наиболее сложный отдел головного мозга. Формирование его в эволюции различных групп позвоночных шло двумя принципиально разными путями, в связи с чем можно выделить два типа конечного мозга: инвертированный и эвертированный (рис. 4). Характерная черта строения инвертированного конечного мозга - наличие в полушариях латеральных, или боковых, желудочков. Такого типа мозг имеют круглоротые, хрящевые и двоякодышащие рыбы, амфибии, рептилии, птицы и млекопитающие-Полушария эвертированного типа лишены желудочков. Единственной группой позвоночных с таким типом конечного мозга; являются лучеперые рыбы, Actmopterygii.
В конечном мозге выделяют лежащие дорсально паллиальные и расположенные вентрально субпаллиальные отделы. Первые представлены кортикальными формациями, вторые -
ядрами конечного мозга. Подкорковые образования конечного мозга так же, как и кора, проходят сложный путь эволюции и: имеют в своем составе филогенетически разные отделы: палеостриатум, архистриатум и неостриатум.
Таким образом, у всех позвоночных формируется сходный тип строения мозга. Однако следует еще раз подчеркнуть, что мозг позвоночных, претерпев длительную эволюцию, у современных их представителей обнаруживает удивительное разнообразие в степени развития одноименных отделов (рис. 5, 6).
Показано, что существует определенная зависимость между массой мозга, массой тела животного и уровнем развития его - ЦНС. Эту зависимость выражают двумя коэффициентами: коэффициентом энцефализации (k) и коэффициентом аллометрииг (α). Они вычисляются по формуле Е = Kpα или чаще в логарифмической форме: lgE = lgk + αlgР, где Е - масса мозга, Р - масса тела. Для каждого конкретного случая k влияет на угол наклона аллометрической кривой, определяя ее значение.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|