- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Борис Ефимович Мельник Мареш Самойлович Кахана 7 страница
Мы считаем, что ергические системы гипоталаму<] участвуют в регуляции ' гомеостаза эндокринных, oj менных и нейровисцеральных процессов. Особое значение эти системы имеют в передаче импульсов по нисходящим нервным волокнам в различные этажи среднего, продолговатого и спинного мозга. Как выявлено с помощью электрофизиологического метода, изменение гомеостаза приводит к передаче нервных импульсовД к лимбическим структурам, что осуществляется чере восходящую систему ергических структур. Не исключ но, что миндалина и гиппокамп играют роль моду/ тора через ту же систему. Особое значение в проце сах стресса имеет мамиллярный комплекс, имея щий нисходящие связи с надпочечниками и восходи щие (через форпике и перегородку) с лимбическй^ структурами. Возбуждение мамилляриого компле са можно сравнить с «зажиганием мотора», в дальней шем уровень реакции зависит от качества и количества катехоламинов. Не исключено, что дофамииергическа" система при участии стриарных образований обеспеч вает основные двигательные рефлексы организма бё выражения эмоций. Норадреналиновая система вклв чает «зоны удовольствия» — положительные эмоции, адреналиновая — в основном отрицательные.
Но самые сложные процессы в гипоталамусе происходят в результате биохимических превращений, приводящих к синтезу разнообразных гормонов, выполняющих специфические функции. Биохимические компоненты гипоталамуса и их связь с обменным процессом в
различных нейронах (в особенности с пейросекрецией И специфическими функциями отдельных центров) мало изучены. Несмотря на то, что гидратация нервной системы очень стабильна, уместно напомнить, что по сравнению с другими тканями количество воды в ней наибольшее. Колебание гидратации ткани мозга невысокое, однако К. И. Пархон, М. С. Кахана и В. Мырза (1929) показали, что баланс воды может быть изменен различными гормонами. Так, гидратация головного мозга у морских свинок достигает 80,5'%, занимая второе место после гидратации семенников, наиболее гидрати-рованных среди всех органов (85,9%).
Из проведенных опытов выявлено, что изменения в гидратации головного мозга колеблются в пределах + 0,68% —0,29'%. Уменьшение гидратации мозга наблюдалось после удаления надпочечников, щитовидной железы, околощитовидиых желез, яичников, семенников, после введения гормонов коры надпочечников, семенников, яичников, нейрогипофиза (вазопрессин, оксито-цнн) у беременных женщин; повышение гидратации тканей мозга выявлялось после гипертиреоидизации,ги-пертимизации, введения плацентарных препаратов, адреналина.
Мало сведений имеется о роли ионного состава в функциях гипоталамуса. Известно, что Л. С. Штерн с соавт. (1946), М. С. Кахана (1947) и др. показали, что пнтравентрикулярное введение солей кальция вызывает торможение в центрах гипоталамуса, приводящее к сонному состоянию (вероятно, блокирование кальцийре-денторов выключает адрепергическне структуры), и, наоборот, введение солей калия вызывает возбуждение. К. И. Пархон, М. С. Кахана (1929) наблюдали уменьшение содержания кальция в тканях мозга после удаления околощитовидных желез. Введение парат-гормо-на дает противоположный эффект. Известна роль калия и натрия, влияющих на мембраны ергических нейронов, которая наблюдается и в гипоталамусе. Hag-gendal (1970) и др. выявили, что особо важную роль играют ионы Са++. Они способствуют выходу норадрс-Налина из пресинаптнческой мембраны. Приток ионов Си++ в нервную термииаль останавливает выделение медиатора. От концентрации Mg++, Fe++, Fe+++ зависит поглощение медиаторов гранулами в пресинапти-
ческой мембране. Интересные данные имеются о ролц Mg++, Ca++, AI+++, которые увеличивали активно^ декарбоксилазы (Christenson ct al., 1971) Активное! катехол-О-метилтрансферазы зависит от присутств? Mg++ в среде, хотя Мп++, Са++, С1_ могут его заи нить. В биохимических процессах активно участвую^ фосфор, фосфатные соединения, регулирующие щелочной резерв й входящие в состав АМФ, АДФ, ЛТФ, фос-фолипидов.
Мало изучена роль микроэлементов (J, Zn, Си других) в обменных процессах гипоталамуса.
К. И. Погодаев (1970—1980) показал, что в ochoij стресса лежит молекулярный механизм, протекающн на уровне органоидов клеток, среди которых ведущая роль принадлежит метаболической структуре митохок дрий. После бега или плавания в митохондриях нейг нов мозга наблюдается максимальное сопряжение дь хания с фосфорилированием, сопровождающееся более высоким потреблением кислорода и осмотической кое центрацией калия в матриксе этих органелл. При эте снижается потенциал действия и облегчается прохож/ ние нервного импульса. При утомлении происходи деэнергизация митохондрий, снижается скорость, коэс] фициент фосфорилирования и утилизация кислорода может понижаться осмотическая компартментализация калия. После электрошока в системе нейрон—нейро^ глия глия получает К+ и теряет Na+, тогда как в не?'" ронах содержание К+ снижается, a Na+ повышается Количество К+- и Na-АТФ-азы не изменяется или cm жаетея.
Гипоксия и другие процессы вызывают у потомства крыс повышенную судорожную активность, нарушен* энергетического обмена в нейронах, связанные со снижением воспроизводства АТФ. Профилактически эмбрионам можно ввести глюкозу, АТФ, сукцинат калия глутамат и метионин для обеспечения пластической обмена.
Углеводный обмен в гипоталамусе представляет са бой особый интерес, поскольку в вентромедиальнв ядрах имеются гликорецепторы, воспринимающие коле бания гликемии, вызывающие чувство голода или насыщения. Вообще все нервные клетки чувствительны понижению глюкозы в крови; с другой стороны, в нк
мало гликогена. Он находится в связанном и свободном состоянии. В наших опытах под влиянием инсулина количество свободного и связанного гликогена в передней части гипоталамуса повышалось значительно, в задней части — менее выражение После введения аминазина количество гликогена снижалось и в передней и в задней частях гипоталамуса. Инсулин восстанавливал запасы гликогена, аминазин, наоборот, уменьшал.
При стрессе, вызванном многократными оборонительными реакциями, количество гликогена как в передней, так и в задней частях гипоталамуса уменьшается. Распад гликогена происходит под влиянием циклического аденазинмонофосфата (АМФ). По мнению Sutherland et al. (1970), АМФ является рецептором для биологических аминов при их физиологическом действии на эффекторные клетки, β-адренсргические блока-торы могут препятствовать этому воздействию. АМФ принимает участие как в медиаторных, так и в гормональных функциях биогенных аминов. АМФ в гипоталамусе также активизирует АТФ, который стимулирует выход аминов из везикул пресинаптических структур. С другой стороны, фенотиазины, пурины, ионы кальция блокируют синтез АМФ.
Липидный обмен в центральной нервной системе и особенно в гипоталамусе изучен недостаточно. Так, по данным Yatsu, Moss (1972), фракция митохондрий головного мозга крыс способна катализировать удлинение линолевой и линоленовой кислот до С22—Сг3 соответственно в присутствии НАДФ-Н, НАД-Н, АТФ и 14С-ацетил КоА. В опытах с растворимыми ферментами различных отделов мозга показано, что в жирные кислоты включаются две С-единнцы при участии мало-нил-КоА.
Akino, Shimajo (1970) выявили, что в мозге куриного эмбриона происходит синтез фосфатидил-инозитидов, в котором участвуют липонуклеотидцитидин и дифос-фат-диглицерид. В мозге цыплят найден фермент, катализирующий ацилирование сфингозина тиоэфирами КоА до образования церамида, то есть ацилефингозина. М. И. Прохорова с соавт. (1972) изолировали из мозга крыс 8 фракций ганглиозидов. Dicessare, Dain (1971), изучая активность трансферазы, катализирующей Х-ацетил-галактазамин на ганглиозид, показали, что
большая часть ее локализовалась в микросомах (441 и митохондриях (З6'%) головного мозга крыс, приче! в онтогенезе активность ее снижалась. По данным Д-. се et al. (1971), субклеточные частицы мозга способны к синтезу ганглиозидов с использованием эндоге акцепторов, причем каждый из них (моно-, ди- и три-сиалоганглиозиды) синтезируются независимо от дру. гих. Изучение распределения жирных кислот и основа ний в длинной цепи ганглиозидов головного мозга пс звоночных позволило Avrova и Zallelinsky (1971 сделать вывод о появлении у млекопитающих (в отл* чие от низших позвоночных) самостоятельного синтез ганглиозидов. Возможные пути синтеза ганглиозидов | других сфиигогликолипидов изучены Eichberg et al (1969).
В литературе имеется ряд работ об обмене фосфо; пидов в тканях нервной системы в процессе миелвд зации. При заболеваниях нервной системы изучены фосфолиниды миелина (Е. М. Крепе, 1967; Д. А. Чев вериков, 1967; И. М. Прохорова с соавт., 1972).
Как показали Smith и Mayer (1911 —1912), холестЯ рин является важнейшим компонентом нервной ткани, где он составляет около 20% липидов. Наибольшее кМ личество холестерина находится в свободном состоянии, это характерно для взрослых животных. На ранних стадиях эмбриогенеза в мозге человека и живот-пых имеется эстерифицированный холестерин. Eichberg el al. (1969) наблюдали в раннем возрасте у детей в период миелинизации увеличение содержания эфиров холестерина до 20%. Значительное количество эфиров холестерина переходит в спинномозговую жидкость, сщ ставляя 50'% от общего количества холестерина в ней (0,26 мг на 100 мл). В числе основных жирных кислот в эфирах холестерина мозга находили олеиновую, пальмитиновую и пальмитилолеиновую, а потом после завершения миелинизации — арахидоиовую.
Smith,Mayer (1972) показали наличие эстернфици-рующих и гидролизующих ферментов в головном мозге взрослых животных. Согласно авторам, холестеролэсиИ раза и гидролазы находятся в митохондриях и микросомах. Их активность повышается с возрастом, параллельно процессу миелинизации. Эфиры холестерина на* капливаются в нервной системе до 25% от общего
НО
количества холестерина в результате дегенерации. Chc-valier, Lutten (1971) считают, что синтез холестерина происходит во время образования и обновления нервных клеток, причем для него характерен замедленный темп. Пути и скорости синтеза холестерина зависят не только от вида и функциональной активности ткани, но л от индуцирующего влияния патологического процесса.
Согласно М. С. Кахана и Л. К- Патуловой (1962), у кроликов с возрастом (от 8 до 200 дней) увеличивается содержание фосфолипидов в передней и задней частях гипоталамуса. У взрослых кроликов введение адреналина увеличивает количество фосфолипидов в гипоталамусе; аминазина — несколько его уменьшает. Содержание холестерина в гипоталамической области больше, чем в коре мозга; в передней части гипоталамуса больше, чем в задней. После введения аминазина содержание холестерина в гипоталамусе уменьшается, причем более значительно в его задней части. Явления стресса, вызываемые многократными оборонительными рефлексами, уменьшают количество холестерина как в передней, так и в задней частях гипоталамуса.
Недостаточно исследован в гипоталамусе обмен нуклеиновых кислот и белков. М. С. Кахана и Л. К. Па-тулова (1962) обнаружили, что с возрастом (от 8 до 200 дней) у кроликов количество ДНК и РНК в гипоталамусе уменьшается. Более выраженное уменьшение наблюдается у 6-месячных кроликов и в передней и в задней частях гипоталамуса. По Jalima (1966), с возрастом в гипоталамусе уменьшается количество всех видов РНК, но особенно информационной. Отношение
•------ в РНК взрослых животных меньше, чем в РНК
молодых.
М. С. Кахана и 3. В. Гандзюк (1961) наблюдали в онтогенезе (с 12 дней до 2 мес) у кроликов увеличение общего количества белков, в дальнейшем (до 12 мес) — тенденцию к его уменьшению. Такие изменения происходят как в передней, так и в задней частях гипоталамуса. Что касается фракций белков передней части гипоталамуса, то с возрастом количество альбуминов повышается, содержание альфа и гамма-глобулинов возрастает, бета-глобулинов снижается. В задней части гипоталамуса количество альбуминов несколько повы-
шается, альфа и бета-глобулинов незначительно увеличивается (до месяца), а затем уменьшается, содержу ние гамма-глобулинов снижается (до месяца), а затем повышается. В гипоталамусе мало альбуминов и больше глобулинов, чем в других тканях.
При стрессовых реакциях, вызванных многократными оборонительными реакциями, общее количество белка в передней и задней частях гипоталамуса уменьша-ется. Введение аминазина снижает содержание общего белка в обеих частях гипоталамуса. Уменьшается фракция альбуминов, бета- и гамма-глобулинов; меньше изменяются альфа-глобулины. Под влиянием инсулина возрастает количество общего белка в передней и задней частях гипоталамуса. Фракция альбуминов и гамма-глобулинов увеличивается, количество альфа- и бета-глобулинов уменьшается в обеих частях гипоталамуса.
Р. В. Дорогая с соавт. (1969) изучали фракции белков гипоталамуса методом электрофореза в акриламид-ном геле и выявили 23 фракции. У взрослых кроликов в первой половине гистограммы отмечали увеличение фракции белков. У молодых много свободных (цистин, аспарагин, глютаминовая кислота и фенилаланин) и связанных аминокислот (лизин, тирозин, метионин, вф лин). Среди аминокислот интерес представляет цисте-ин, носитель SH-rpynn. SH-группа в гипоталамусе имеет особое значение, поскольку она входит в состав ферментных систем, кроме того содержат SH-групиы октопептиды и некоторые либерины и статины. С другой стороны, SH-группы входят в состав таких ферментов, как моноаминооксидаза и аденозилметиопин. Fri-edenwald et al. (1942) считают, что моноаминооксидаза содержит SH-группы, обеспечивающие транспорт электронов. Ингибирование этого фермента предотвращается введением глутатиона и цистеина, которые восстанавливают SH-группы (Lagnada et al., 1956). Axel-rod (1957), Blashko (1968), Weinshilboim (1972) и др. указывают на роль SH-аденозилметионина в качестве донатора метилыюй группы КОМТ, который осуществляет О-метилирование аминов и их инактивацию в синаптической щели или в эффекторной клетке. В связи с тем, что ряд тиоловых реагентов (P-CL-7-меркурив бензоат, иодацетамид и др.) являются ингибиторами
-екарбоксилазы, возможно участие SH-групп в активности коэнзимпиридоксил фосфат-декарбоксилазы (Christensen, 1970). В исследовании функции гипоталамуса мы придавали особое значение обмену SH-rpynn. для этого использовали полярографический и гистохимический методы (по Barnet). Из наших данных следует, что количество SH-rpynn меньше в задней части „' больше в передней части гипоталамуса. Это можно объяснить тем, что в передних ядрах цистеин используется для синтеза октопептидов. На содержание SH-rpynn влияют различные гормоны, связанные с их обратным действием на гипоталамус. Так, М. С. Каха-на и Е. Н. Рапопорт (1967) показали, что с возрастом количество SH-групп в гипоталамусе увеличивается. Зведение тиреоидных гормонов вызывает увеличение количества SH-групп у молодых кроликов, а у взрослых (3—4-месячных) — уменьшение. Гистохимическим методом Barnet выявлено, что в цитоплазме нейронов супраоптических и паравентрикулярных ядер меньше SH-групп, в нейросекрете, в межклеточных пространствах — больше, что можно объяснить усиленной активностью нервных клеток у молодых кроликов. Наоборот, у взрослых животных SH-групп больше в цитоплазме нейронов, меньше в межклеточном пространстве, что указывает на снижение интенсивности нейросекре-:ши (А. Таликина, 1970).
Введение фолликулина и прогестерона двухлетним кроликам уменьшает количество SH-групп в нервных клетках указанных ядер по той же причине. У взрослых кроликов под влиянием половых гормонов больше накапливается SH-групп в цитоплазме. Не исключено, что такие изменения имеют отношение к синтезу октопептидов и тиреолиберина.
М. С. Кахана, А. С. Гудима (1973) показали, что у крыс билатеральная адреналэктомия снижает концентрацию SH-групп в передней части и несколько повышает в задней части гипоталамуса. После введения гидрокортизона восстанавливается прежний уровень SH-трупп. Как солевое, так и безводное питание и особенно введение гипертонического раствора хлористого натрия вызывают снижение количества SH-групп в переднем и заднем отделах гипоталамуса. Известно, что гИпогидратация крови усиливает выделения вазопрес-
8 Заказ № 457. ИЗ
сина, антидиуретическое действие которого уменьщае выделение воды из организма. Поэтому усиление син. теза вазопрессина уменьшает количество SH-rpynn гипоталамусе.
Можно полагать, что влияние гормонов на динами. ку SH-групп в гипоталамусе осуществляется через ер. гические структуры гипоталамуса. У ложноадрсналэк-томированных крыс (стрессовое действие) наблюдалось уменьшение количества SH-групн в передней и увели-чение в задней частях гипоталамуса. АКТГ, введенный крысам в дозе 4 ед. на 100 г веса, вызывает снижение количества SH-групп как в передней, так и в задней частях гипоталамуса. А. Я. Местечкина (1968) наблюдала уменьшение количества SH-групп в различных отделах головного мозга после введения кортикостерои-дов. Эти гормоны имеют отношение к серотопинерги-ческим структурам.
Представляет интерес влияние ацетилхолина и хо-лннергических факторов на количество SH-групп в гипоталамусе. Введение ацетилхолина крысам увеличивало количество SH-групп (М. С Кахаиа, Н. К. Токарева, 1975). Уместно напомнить, что А. А. Головин (1963) усиливал нейросекрецию супраоптических ядер при помощи ацетилхолина. Martini (1954) считал, что аце-тилхолин стимулирует синтез вазопрессина, который действует на выделение АКТГ. После введения адреналина крысам в гипоталамической области отмечается увеличение количества SH-групп. Не исключено, что такие изменения связаны с активацией моноаминоокси-дазы и SH-аденозилметионина.
Введение норадреналина крысам уменьшает содержание SH-групп в гипоталамусе. Тиреоидные гормоны имеют такой же эффект. Под влиянием аминазина количество SH-групп в гипоталамусе также уменьшается, что можно объяснить снижением выделения адреналина. Введение серотонина крысам приводит к нарастанию количества SH-групп. Интересно отметить, что адреналин и серотоиин вызывают противоположные эффекты по сравнению с норадреналином и адреналином.
Методы определения SH-групп в гипоталамусе по существу равноценны методам определения количества цистеина. Необходимо подчеркнуть, что цистеин И цистин входят в состав октопептидов и гормонов гипо-
.дамуса. Таким образом, по количеству SH-rpynn |0}КНО судить об активности некоторых ферментов, содержащих эти радикалы. С другой стороны, по количеству SH-rpynn можно судить об уровне образования S— с.групп, об интенсивности синтеза октопептидов и не-к0торых гормонов гипоталамуса (В. Ф. Чернокан, 1980).
Klegg and Klegg (1960) приводят убедительные опыты, показывающие, что меченый цистеин, введенный я гипоталамус, через некоторый интервал времени выявляется в задней доле гипофиза в составе октопептидов. Safran, Sayers (1960) утверждают, что октопептиды влияют на синтез кортиколиберина, химическая структура которого близка к вазопрессину.
Вероятно, интенсивность синтеза октопептидов в гипоталамусе имеет и другое значение. Так, до открытия кортиколиберина и других гормонов, по данным Sawyer (1960) и др.", считалось, что вазопрессин является стимулятором синтеза АК.ТГ.
Есть основания думать, что октопептиды служат материалом, из которого синтезируются некоторые ли-берины и статины гипоталамуса. Так, в химической структуре кортиколиберина и соматостатина имеются S-S-группы и фрагмент октопептидов. В меланоцит ли-берина входят две молекулы цистеина. Октопептиды могут играть роль модуляторов в нервных и гормональных процессах, протекающих в "гипоталамусе.
Кроме того, по данным М. С. Кахана, П. И. Почтарь (1974) и др., после кастрации, при гипотиреозе, зведении дексаметазона и других кортикостероидов количество адреналина и норадреналина в гипоталамусе повышается. Можно полагать, что катехоламины в Дальнейшем влияют на синтез гонадолиберинов, тирео-•тиберина и кортиколиберина.
Не исключено, что адреналин кроме медиаторной Функции стимулирует адренергические системы, влияющие нисходящим путем на мозговую часть надпочечников, участвуя тем самым в различных обменных процессах (гликогенолиз, гликолиз, липолиз, расщепление белков). Расщепление липотропина, АКТГ, МСГ приводит к образованию эндорфинов, энкефалинов и нейро-"еитидов, увеличению количества цистеина в гипоталамусе; появляются и другие аминокислоты — глютами-
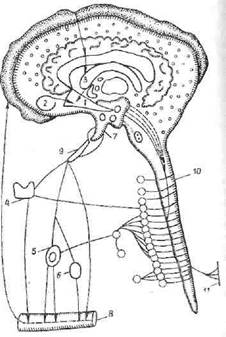
Рис. 12. Нейрогуморальные и гормональные компоненты стресса:
1 — гипоталамус (синтез либеринов, статинов, эндор-финов, энкефалинов); 2 — миндалина (влияние на ад-ренергические структуры гипоталамуса); 3 — связь с корой мозга, гнппокампом (холин- и серотонинерги-ческие структуры гипоталамуса); 4 — щитовидная железа (влияние тиреотропного гормона. выделение в кровь гормонов Т3, Тч); 5 — АКТГ, надпочечники (выделение кортикостероидов, катехоламинов); 6 — половые железы, панкреас (выделение инсулина, глюкаго-на); 7 — задняя доля гипофиза (выделение октопепти-дов); 8 — кровь и обратное действие гормонов на структуры мозга; 9 — сосудистая система (выделение гормонов гипофиза); 10 — ретикулярно-симпатичеекзя система; 11 — болевые рецепторы (гистамин. кинины. нростагландины)
новая кислота, аргинин, фенилаланин, из которых синтезируются субстанция Р, опиоидные белки, либерины „ статины (рис. 12).
На повышение количества цистеина в гипоталамусе влияют ацетилхолин и серотонин, активизируются и различные ферменты из группы моноаминоксидаз.
Норадреналин и тиреоидные гормоны уменьшают количество SH-rpynn, способствуют образованию S-S-групп, входящих в состав октопептидов, кортиколнберина, соматостатина. В свою очередь тиреолиберин влияет на синтез пролактолиберина, пролактостатина, увеличивая выделение пролактина. Таким образом, ок-топептиды являются не только сырьем, но и стимулятором синтеза кортиколнберина, АКТГ и иейропептидов.
9. СТРЕСС И НЕВРОЗЫ
Как известно, различают три типа экспериментальных неврозов: а) с преобладанием процессов возбуж. дения; б) с преобладанием процессов торможения' в) циркулярный, при котором указанные состояния проявляются последовательно.
Согласно данным К. М. Быкова и И. Т. Курцина (1947), при нарушении корковых функций во взаимоотношениях между корой мозга и подкорковыми образованиями наступают определенные изменения. Под влиянием сильных воздействий, вызывающих нарушение высшей нервной деятельности, в первой фазе в коре головного мозга образуется очаг возбуждения; в дальнейшем, если продолжается вредное воздействие на центральную нервную систему, наступает охранительное торможение коры головного мозга. Б результате этих изменений возникает положительная индукция подкорки, в частности гипоталамических центров. Указанные центры начинают работать без полноценной регуляции коры мозга, что приводит к различным патологическим явлениям со стороны функции внутренних органов. Крртико-висцеральная патология наблюдается при неврозах, но тем не менее промежуточные механизмы, приводящие к нарушениям функций различных систем, полностью не выявлены. Неврозы вызывают различные стрессы.
В. В. Суворова (1975) считает, что основные виды стрессовых состояний у человека следующие: социальный, психический и физиологический. Согласно автору, в стрессе важную роль играют эмоции и утомление. В процессах стресса участвуют вегетативный отдел нервной системы и гормоны (адреналин, кортикостероиды, инсулин). Выделение гормонов регулируется гипотала-мо-гипофизарной системой с помощью АКТГ. Выявле-
на
'
b! электроэнцефалографические показатели общего адаптационного синдрома. Повышение активности тета-pii-гма характеризует стрессовый этап, связанный с максимальной синтетической активностью мозга. Доминирование парасимпатической активности сопровождается Усилением активности дельта-ритма. Высокоамплитуд-нэя, нерегулярная дельта-синусоида становится господствующей при активации переднего гипоталамуса и лреоптических ядер у животных. Доминирование парасимпатической активности переднего гипоталамуса при подавлении коры мозга и подкорковых структур имеет типичное поведенческое выражение. У животных наблюдается оцепенение с уменьшением сенсорной и двигательной активности. Такое состояние характерно для церебрального стресса первосигнального типа при сры-зе высшей нервной деятельности в сторону торможения. Для периферического и психического второсигнального стресса оцепенение не характерно. Нарушение вегетативного реагирования в сторону преобладания симпатического или парасимпатического регулирования определяет характер дезорганизации деятельности и поведения при стрессе.
В течение 1952—1959 гг. нам удалось более подробно исследовать некоторые изменения в гипоталамусе и функциях желез внутренней секреции, возникающие при экспериментальном стрессе.
У различных видов животных (кроликов, собак, кошек) был вызван срыв высшей нервной деятельности в сторону возбуждения. Более выраженные изменен яя были отмечены у кроликов, у которых выработались двигательно-пищевые условные рефлексы. После многократных оборонительных реакций у подопытных животных (по сравнению с контрольными) угасали выработанные условные рефлексы, ориентировочная реакция исчезла с трудом, что указывало на развитие процесса торможения в коре головного мозга. Одновременно были выявлены определенные изменения биоэлектрической активности центральной нервной системы по сравнению с канальным фоном и картиной ЗЭГ у контрольных животных. Оказалось, что биотоки коры мозга уменьшились по частоте и амплитуде, вследствие чего преобладали медленные колебания; в то же время в гипоталамиче-ской области процессы возбуждения нарастали. Эти
данные явились подтверждением того, что при эксперт ментальном неврозе у кроликов развивались процесск, торможения в коре мозга и возбуждения в гипоталаму се. У этих животных был отмечен ряд изменений со сто роны желез внутренней секреции, повышение активно, сти щитовидной железы, усиление выделения адренали-на, вазопрессина, антидиуретического и адренокорт,,. котропного гормонов с некоторой активацией коры надпочечников. В то же время уменьшалось выделение гонадотропного гормона, инсулина; половая функция была подавлена.
В связи с этими данными небезынтересно напомнить, что у пойманных зайцев (Stern et al., 1958), особенно под влиянием лая собаки, возникают специфические изменения в нервных клетках гипоталамуса, указывающие на усиление их секреторной деятельности. Наиболее выраженные изменения наступают преимущественно в нервных клетках супраоптических ядер —. увеличивается объем клеток, отмечается гипертрофия ядер и ядрышек. Авторы считают, что при этом усиливается синтез протеинов в клетках и мобилизуется се-ротонин.
Как показали другие опыты, не во всех зонах гипоталамуса функции усилены. Это было выявлено при изучении биоэлектрической активности ядер передней и задней частей и других зон гипоталамуса. Выяснилось, что биоэлектрическая активность в центрах задней части гипоталамуса высокая, а в других зонах— несколько подавлена. У животных также были отмечены снижение аппетита, понижение безусловного слюноотделения и уменьшение диуреза. У собак и кроликов при экспериментальных неврозах одни центры гипоталамуса возбуждаются, а другие угнетаются. Это значит, что положительная индукция подкорки не охватывает гипоталамическую область в целом.
Исходя из вышеизложенного, большой интерес представляет исследование изменений центральной нервной системы и функций желез внутренней секреции у больных, страдающих неврозом. Из наших наблюдений следует, что у больных истерией, психастенией и неврастенией возникают определенные изменения в функциях гипоталамуса. Так, у больных истерией и психастенией преобладает возбуждение адренергических структур ги*
ш
\ротала^уса; некоторое усиление функций щитовидной ^елезы,. медуллярной части надпочечников и задней доли гипофиза. У неврастеников и некоторых психастеников после короткого возбуждения адренергических структур гипоталамуса наступает подавление функции некоторых нервных центров этого отдела. Это приводит чзсто к астении, подавленности, сонливости, потере трудоспособности.
Основная роль в возникновении различных эмоциональных процессов принадлежит коре мозга, а затем лпмбическим образованиям, в частности гипоталамиче-ским центрам. Гипоталамическая область является ключевым аппаратом эмоциональной жизни, одновременно связанным с корой мозга и эндокринной и вегетативной
I системами. Половые рефлексы и инстинкты продолжения рода тесно связаны с функциями преоптической зоны и аркуатного ядра гипоталамуса. Различные защитные реакции и другие эмоциональные переживания осуществляются через симпатическую вегетативную и эндокринную системы. При этом наблюдается ряд фи-
I зиологических явлений: тахикардия, изменение кровяного давления, сокращение гладкой мускулатуры кишечника и мочевого пузыря, усиление потоотделения н т. д., которые зависят от вегетативных центров гипоталамуса.
Можно считать, что психическая травма сопровож-
| дается настоящими кортико-диэнцефаловегетативными рефлексами. Нередко условные рефлексы усиливают или ослабляют безусловные. Как было отмечено рядом исследователей, иногда условные рефлексы сильнее безусловных и тормозят последние.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|