- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Борис Ефимович Мельник Мареш Самойлович Кахана 2 страница
Вместе с тем границы между этими категориями гомеостаза условные — в физиологических стрессах наблюдаются гормональные, обменные и нейровегетативные компоненты.
6.1. ТЕМПЕРАТУРНЫЙ СТРЕСС
6.1.1. Гипотермический стресс
Различают холодовой и тепловой стрессы. Гипотермический стресс у животных обычно протекает сезон-но. Применяется он и в хирургических операциях на сердце. В основном охлажденная кровь влияет на медиальные и мамиллярные центры гипоталамуса. При этом повышается тонус симпатической нервной системы, усиливается выделение катехоламинов и тиреоид-чых гормонов, стимулируются окислительные процессы, гликогенолиз, липолиз, гликолиз. Посредством химической терморегуляции сохраняется относительное Постоянство температуры тела (рис. 2).
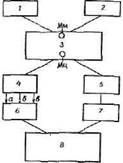
Рис. 2. Схема стрессорного воздействия холода на организм:
1 — холодная кровь; 2 — хо-лодовые рецепторы, таламо-мамиллярный пучок; 3 — гипоталамус. Мм — мамиллярные центры, Мц — медиальные центры; 4 — ретикулярная формация, симпатическая система; 5 — миндалина (поведение. Защитная реакция, отрицательная эмоция); 6 — кровь: а — катехоламнны, б — тиреоид-ныегормоны, в — глюкагон (гликогенолиз. липолиз); 7 — кора мозга (условные рефлексы избегания холода, приема пищи); 8 — восстановление исходной температуры тела (гомеостаз терморегуляции)
В меньшей степени щ процессе поддержания температурного гомсостазиса при гипотермии участвуют физическая терморегуля-1 ция, вазоконстрикция, изменение дыхания, излуче-: иие и т. д. Гипотермический стресс изучается экспериментально для выявления] тонких и сложных нервных и нейрогуморальных процессов. Так, мы показали, что при охлаждении гипо-j таламических центров после субокципитального введения охлажденной спинномозговой жидкости наступает де-j синхронизация ритма в яд-' pax медиальной и мамил-лярной частей. При нара-j стании охлаждения импуль-| сы распространяются на миндалевидные ядра, вызы{ вая мотивационно-поведен-ческие реакции.
Гипотермическая реаю! ция сопровождается уси-1 лепным выделением адре{ налииа, норадреналина, ти] реоидных гормонов, серото-j нина. Изменяются функции медиальной эминенции гипоталамуса, при этом в про-, цесс включаются холинергические, адренергические й серотопинреактивные структуры. Так, Э. И. Ажипа с соавт. (1973) показали, что холодовые раздражения кожи увеличивают количество ацетилхолина в крови.
По данным Бенетато и Улуиту (1959), ссротонин выделяется из нервной ткани промежуточного мозга. Согласно М. С. Кахама и Г. А. Ника (1960), сильное охлаждение гипоталамуса вызывает гипотермическук} реакцию и состояние, подобное парабиозу. В дальнейшем усиливается выделение АК.ТГ и кортикостероидов|
после чего восстанавливается исходная температура тела.
Е. А. Ильин и Л. С. Вознесенский (1973) у здоровых людей в первой половине года зимовки в Антарктиде отметили, что количество 11-оксикортикостсроидов в крови повышается, а в дальнейшем нормализуется. Это происходит, вероятно, в результате тренировки и приспособления организма к среде. В связи с этим представляет интерес работа И. В. Готлебер и Ю. А. Васильева (1973), акклиматизировавших крыс к холоду. При плавании в воде при температуре 0°С авторы наблюдали адаптацию к физической нагрузке. Приспособление к холоду на 19% увеличивает амидирован-ность белков за счет прочно связанных амидных групп.
К- В. Осташков и Т. Е. Орлов (1976) вызвали стресс при селективной абдоминальной гипотермии у собак. Авторы обнаружили некоторое сгущение крови с возрастанием количества эритроцитов и гемоглобина при замедленном СОЭ и выраженной лейкопении, слегка повышенном содержании СОг в венозной крови.
Т. Г. Гамалий и Л. Г. Гречук (1973) при подобном стрессе наблюдали снижение общего белка крови, γ,-рУ и γ-глобулинов, остальные фракции не изменялись. Резорбция глюкозы в кишечнике снизилась.
3. А. Гершенович с соавт. (1965) показали, что гипотермия влияет на уровень мочевины в мозге. До некоторой степени она помогает животным выжить после стрессорных воздействий. Мочевина синтезируется при повышении активности аргиназы. По данным В. С. Шу-галей и Л. С. Козиной (1973), в первые дни гипотермии понижаются активность аргиназы и уровень мочевины в мозге. В дальнейшем содержание мочевины растет. С другой стороны, авторы считают, что мочевина во взаимодействии с белками регулирует активность аргиназы и других ферментов, способствуя быстрому выходу из состояния стресса.
Гаким образом, гипотермия вызывает катаболиче-скую фазу — выделение катехоламинов и тиреоидных юрмонов, а затем анаболическую фазу — выделение АКТГ и кортикостероидов.
6.1.2. Гипертермический стресс
Перегревание организма считают физиологические, стрессом, который наблюдается у животных и человека Однако резко выраженная гипертермия сопровождает ся парабиозом и протекает так же, как тепловой нар коз. Эволюция теплорегуляции показывает, что внача ле у животных формируются адренергические цеитрь гипоталамуса, обеспечивающие приспособление к холо) ду, а затем лишь — центры передней части гипотала муса, регулирующие явления перегревания. При их аК тивации снижается интенсивность обменных процессов уменьшается выделение катехоламинов и тиреоидны.} гормонов, снижается тонус симпатической и повышает ся тонус парасимпатической нервной системы, расши) ряются сосуды. Как показали наши электрофизиологи ческие исследования, в передних ядрах гипоталамус наблюдается особенно выраженная синхронизация рит] ма, которая охватывает в основном холинергическя структуры мозга. В дальнейшем иррадиация медлег ных ритмов распространяется на миндалевидный kon плекс, гиппокамп, кору мозга и обеспечивает мотива ционно-иоведенческие реакции у животных и человека1 В этих процессах участвуют серотонинреактивные струк туры. Серотонин, по мнению Бенетато и Улуиту (1969) является термолитическим медиатором.
Распределение 5-окситриптамина в нервной систеб позвоночных более или менее соответствует распред< лению симпатина (Vogt, 1954). Его больше в сером в ществе нервной системы, чем в белом, очень много вегетативных центрах и в area postrema. Найден он в обонятельном мозге. Наибольшая его концентраци по мерс убывания обнаруживается в структурах ЦНС следующем порядке: гипоталамус, средний мозг, епш ной мозг, кора мозга. В зрительном нерве и в мозже1 ке 5-окситриптамин не обнаружен (Amin, 1954). 5-0 ситриптамин делает кошек вялыми и сонливыми и Я давляет стрихнинные судороги. Раувольфин и резерпй! действуют, высвобождая из мозга 5-окситриптам1 (Flescher, 1956). Спустя 24 ч после введения этих в ществ из головного мозга кролика исчезают 90% 5-q ситриптамина (Yless et al., 1956). При раздражен центрального конца блуждающего нерва в головне
мозгу также высвобождается 5-окситриптамин. Антагонистом резерпина служит диэтиламид лизергиновой кислоты; его введение уменьшает длительность периодов покоя, наступающих после высвобождения серото-нина (5-окситриптамина), а также вызывает у людей галлюцинации и шизофреническое поведение.
В связи с этим отметим, что С. X. Хайдарлиу, Б. П. Тонкоглас (1980) и др. изучали у крыс количество ацетилхолинэстеразы в гипоталамусе при нагревании в термокамере до температуры 45°С. У контрольных животных активность ацетилхолинэстеразы в переднем гипоталамусе оказалась ниже, чем в заднем на 23%. В первые 70—80 мин наблюдалось усиление двигательной активности и появление судорог. В то же время активность ацетилхолинэстеразы повышалась на 30—46%. Через 100—115 мин изменений ацетилхолинэстеразы не наблюдалось. Эти данные совпадают с нашими электрофизиологическими исследованиями, согласно которым нагревание гипоталамуса приводит к активации холинергических структур. По нашим данным, в паравеитрикулярных ядрах гипоталамуса преобладают холинергические нейроны.
К- В. Осташков и Т. Е. Орлов (1973) после переохлаждения обогревали животных, что вызвало некоторую гипоксемию и гиперкапнию артериальной крови. В результате ослабления дыхания содержание кислорода в венозной крови возрастало на 27%. В. В. Бонда-ренко с соавт. (1973) у людей, подвергнутых отрицательной аэроиоиизации, под влиянием тепловых воздействий наблюдали меньшие изменения температуры тела и кожи, легочной вентиляции, частоты пульса и артериального давления. В. Н. Васильков (1973) наблюдал, что механизмы терморегуляции у забойщиков шахт, работающих при температуре до 30°С, оказываются в состоянии большого напряжения, теплопродукция у них составляет 2500 кал, пульс — 142 удара в мин, температура тела повышается на 0,8°С, дефицит веса тела — <*>' кг, производительность труда снижается.
Таким образом, небольшое нагревание можно считать антистрессовым воздействием, резкое и длительное -_ стрессовым.
6.2. ПИЩЕВОЙ СТРЕСС
6.2.1. Гипогликемический стресс
При голодании возникают отрицательные эмоции при насыщении — положительные. Современные элек трофизиологические исследования показали, что голо дание возникает в результате гипогликемии. В наши; опытах уменьшение количества глюкозы в крови влия ло на гликорецепторы вентромедиальных ядер (аффе рентные). Нейроны этих ядер полисенсорные. Они pea, гируют на глюкозу, на афферентную импульсацию и; полости желудка и других отделов желудочно-кишеч] ного тракта и на электрические раздражения (Olds 1958; М. С. Кахана с соавт., 1961). В случае гипогли кемии в вентромедиальных ядрах обнаружили десин; .хронизацию ритма (возбуждения), которая передава лась к вентролатеральным ядрам (эфферентным). Эт] ядра через адренергические центры ретикулярной фор мации вызывают выделение катехоламипов и повыша ют тонус симпатической нервной системы, усиливаю щей выделение тиреоидных гормонов, в свою очеред! увеличивающих скорость окислительных процессов, гли когенолиза, липолиза. В связи с нарастанием гипогли кемии десинхронизация ритма центров гипоталамуса] распространяется на миндалевидные ядра, возникаю.! мотивационно-повсденческие реакции, характерные дл хищников (гнев, агрессия). У животных наблюдает^ физиологический голодный стресс, протекающий в за висимости от их видовых особенностей. У человека г погликемический стресс входит в ритм функции орг низма. Нарушение этого ритма (десинхропоз) вызыв ет иногда язву желудка, дистонию и т. д. (рис. 3 Некоторые авторы считают, что длительный десинх^ ноз сокращает продолжительность жизни животны (А. Войкулеску, 1972; Б. С. Лякритский, 1973). Эк] периментальныи пищевой стресс наблюдается у жива ных и в результате несовпадения поиска и поведепче ского акта с моделью будущих событий, акцепторе действия, например, с отсутствием пищи в кормуш! (П. К. Анохин, 1964; К- В. Судаков, 1972).
Следовательно, при голоде развертывается первг фаза стресса — выделение катехоламинов и тиреои
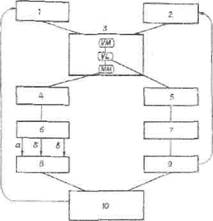
Рис. 3. Схема стрессорного поздснствня голода: I — гипогликемия ;2 — голодные сокращения желудка; 3 —гипоталамус: VM — вентромеднальиые ядра, VL — вентролатеральные ядра, ММ мамнллярные
ядра; -4 — ретикулярная формация; 5 — миндалина (поведенческие, двигательные, пищевые рефлексы); 6 — симпатическая нервная система: а — выделение, катехоламииов, 6 — тиреоидин* гормонов, в — глюкагона; 7 — кора мозга (пищедобыва-тельные рефлексы); 8 — печень (гликогенолнз); 9 — прием пищи (вода, углеводы, жиры, белки); 10 — кровь (восстановление гликемии)
ных гормонов. В дальнейшем выделенные кортикосте-роиды подготавливают органы пищеварения к приему пищи. Если пища не поступает, развивается язва желудка. Прием пищи прекращает стрессовую реакцию и предупреждает возникновение язвы.
6.2.2. Гипергликемический стресс
После приема пищи отрицательные эмоции исчезают и появляется чувство насыщения, удовлетворения потребности организма, восстянявливается гомеостаз углеводного обмена. Насыщение организма приводит к
3 Заказ № 457.
гипергликемии. Б то же время выделение инсулин^ АКТГ, кортикостероидов повышается — реакция наш; минает второй период стресса.
Электрофизиологические исследования выявили сЯ хронизацию ритма в гипоталамусе, миндалинах и гЩ покампе. Из данных Brobeck, Anande (1965) следуе] что нарастание количества глюкозы в крови влияет вектромедиалькые центры. Авторы считают, что ме>: ду этими ядрами И вентролатеральными существу* решшрокные взаимоотношения. Однако, согласно на щим данным, после возбуждения или торможения Bemj ромедиальных ядер иррадиация передается к вента латеральным ядрам. При гипергликемии наблюдал торможение этих центров. Липиды, холестерин, амине кислоты так же влияют на биоэлектрическую актиЕ ность вентромеднальных ядер. Одновременно из поло^ ти желудка информация к этим ядрам направляете через блуждающие афферентные волокна, в результа^ чего в них возникают явления синхронизации ритма торможения, распространяющиеся на вентролатералЕ ные ядра. На наш взгляд, эти формации — эфферещ ные. Они повышают тонус парасимпатической нервно| системы, активизируют холинергические формации зону удовлетворения гипоталамуса. Эти зоны xopoi изучены Olds (1959) и другими с помощью самора^ дражения электрическим током. По нашему мнении' они содержат норадреналиновые нейроны, имеющк нервные связи с мозговой частью надпочечников. В мозговой части Hillarp (1960) выявил флюоресцируют* адреналиновые и нефлюоресцирующие норадреналинс вые клетки. Мы полагаем, что указанные клетки имен' непосредственные нервные проводниковые связи с нами удовольствия или зонами отрицательных эмоце гипоталамуса. При страхе, агрессии, гневе выделяете адреналин, при положительных эмоциях — норадренй лин.
Из наших электрофизиологических исследован!; следует, что в гиппокамне также имеются норадреналЕ новые нейроны, ответственные за мотивационно-поЕ денческие реакции, успокоение, сонливость. При нас1 щении синхронизация ритма иррадиируется гиппокамп. Вероятно, что холинреактивные и серот
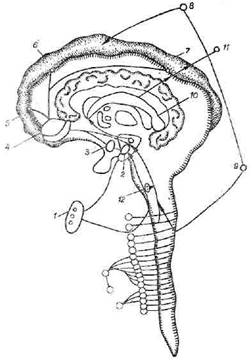
Рис. 4. Лймбико-гипоталамические отношения при голоде и насыщении:
1 — афференгы гипоталамуса (а — гипогликемия, б — голодные сокращения желудка;
2 — вентромедиальные ядра (афферентные) — гликорецелторы;
3 — вснтролатсральные ядра (эфферентные); 4 — миндалина (пищевые поведенческие реакции, агрессивность); 5 — обонятельный анализатор; 6 — слуховой анализатор; 7 — зрительный анализатор; 8 — центр пищедобывательных рефлексов и отрицательных эмоций; 9 — пища (гипергликемия, содержимое желудка, передача импульсов в вентромедиальные ядра, гиппокамп); 10 — гиппокамп (поведенческие реакции, успокоение); 11 — кора мозга (насыщение, положительные эмоции); 12 — центр вагуса
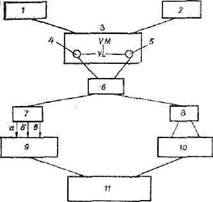
Рис. 5. Схема регуляци гомеостаза глюкозы (алгоритм системы).
1 — гипогликемия; 2 — гипергликемня; 3 — гипоталамус: VM — вентромеднальныс ядра; VL — вснтролатеральные ядра; 4 — адренергические структуры гипоталамуса; 5 — вагоинсулярнмй центр гипоталамуса (кортиколиберин. ЛКТГ); 6 — ретикулярная система (симпатические и парасимпатические нисходящие пути); 7 — симпатическая нервная система; а — выделение катехоламинов. б — ти-реоидных гормонов, в — глюкагона; 8 — парасимпатическая система (инсулин, кортикостероиды); 9 — гликогенолиз; 10 — гликогенсз, неогликогенез. липогенез. 11 — гомео-стаз глюкозы (восстановление исходной гликемии)
нинергические структуры также участвуют в процесс; насыщения (рис. 4, 5).
Следовательно, насыщение, гипергликемия угнетак развитие стресса, однако способствуют развертыванй его второй фазы — выделению АК.ТГ, кортикостеро: дов, инсулина.
6.3. РВОТНЫЙ СТРЕСС
Под влиянием вредных факторов у высших млек| питающих и у человека наблюдаются антиперистальт) ческие реакции — рвотный рефлекс. Тошнота и рвот
возникают при интоксикации, укачивании, неврозах, облитерации кишечника и т. д. Нервно-рефлекторные механизмы полностью не раскрыты. В них участвуют центры продолговатого мозга, мозжечка, гипоталамуса и коры мозга. Считают, что древний отрыгивательный рефлекс у травоядных животных (физиологический) превращается в патологический, когда включаются ак-типеристальтические рефлексы различных отделов желудочно-кишечного тракта. В этих процессах участвуют также нервные вегетативные элементы, неспецифические гуморально-гормональные и гипофизарно-адрена-ловые системы, центры насыщения гипоталамуса. Считают, что эти формации влияют на выделение окси-тоцина, вызывая у грудных детей рвоту.
6.4. ГИДРАТАЦИОННЫЙ СТРЕСС
6.4.1. Гипогидратационный стресс
Уменьшение гидратации организма вызывает жажду, которая сопровождается отрицательными эмоциями. Прием воды снижает эти явления, вызывает удовлетворение, иногда временную гипергидратацию, поскольку постоянство количества воды в организме регулируется механизмами гомеостаза.
Выраженная гипогидратация, гипогидремия порождают не только жажду, но и расстройства обменного характера. Гипогидремию мы получили у животных введением внутривенно 20%-пого хлористого натрия. Наблюдали десинхронизацию ритма в супраоптических и вентромедиальных ядрах гипоталамуса. К этим центрам направляется и информация от осморецепторов крупных сосудов, внутренних органов, но главным фактором в механизме гипогидратационного стресса является действие осмотического давления крови на осмо-рецепторы гипоталамуса (Verney, 1949). Эти воздействия, особенно на уровне супраоптических центров, приводят к выделению вазопрессина, в результате чего проявляется его антидиуретическое действие, реабсорбция первичной мочи нарастает, диурез уменьшается; появляется олигурия. В случае гипогидремии десинхроинза-Дня ритма и возбуждение супраоптических центров распространяется на миндалины и гиппокамп — наблюда-
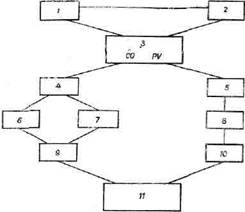
Рис. 6. Схема стрессорного действия жажды (полидипсия) :
1 — гипогидратация крови; 2 — осморецепторы сосудов и органов; 3 — гипоталамус: СО — супраоптические ядра, PV — параоентрнкулярные ядра, осморецепторы гипоталамуса; 4 — задняя доля гипофиза (вазопрессин); 5 — миндалина (поведенческие реакции на жажду); 6 — антидиуретическое и вазопрессорное влияние на сосуды капсулы нефрона; 7 — прессорное возбуждение юкста-гломерулярной ткани, выделение ренина, ангиотензина; 8 — вовлечение анализаторов в поиск воды; 9 — влияние вазопрессина и альдостерона. уменьшение выделения воды; 10 — прием воды; 11 — восстановление гомеостаза водно-солевого обмена (алгоритм гомео-стаза)
ется мотивационно-поведенческая реакция (поиск воды характерная для каждого вида животных. Некотор! авторы считают, что в этих процессах участвуют и ю( стагломерулярные зоны, влияющие на выделение ал| достерона (рис. 6).
Л. В. Серова с соавт. (1973) вызвали дегидратаци! у крыс содержанием их без питья на сухом корме в v чение 3—5 дней. Авторы наблюдали уменьшение вес тела тимуса и селезенки, инволюцию тимуса. В перво фазе стресса отмечается выделение катехоламинов, в; зопрессипа и альдостерона, во второй — кортикост роидов.
6.4.2. Гипергидратационный стресс
Прием воды является антистрессовым фактором. Он восстанавливает водно-солевой гомсостаз — жажда исчезает, наблюдаются положительные эмоции. Однако накопление большого количества воды в организме может вызвать расстройство циркуляции крови и даже отечность. Экспериментально введение больших доз ва-зопрессина или альдостерона приводит к задержке воды в организме и расстройству его функций, поэтому можно говорить о гипергидратационном стрессе. Не исключено, что при этом участвуют холинергнческие и норадренергические нейроны. Установлено, что при гипергидратации уменьшается выделение вазопрессина, временно усиливается выделение воды и, как результат, восстанавливается гомеостаз водного обмена. Выделение кортикостероидов во второй фазе стресса может вызвать отечность.
6.5. РЕСПИРАТОРНЫЙ СТРЕСС
Давно известны явления, которые отмечаются при дыхании в условиях пониженного и повышенного атмосферного давления. Экспериментально можно получить гипоксический, гипероксический, гипобарическнй и гипербарический стрессы.
6.5.1. Гипоксический стресс
По данным И. А. Аршавского с соавт. (1977), в антенатальном периоде важным фактором развития плода является физиологическая гипоксемия.
С момента, когда возникает и созревает скелетная мускулатура и нервные центры, регулирующие ее деятельность, что в основном совпадает с установлением связи эмбриона с материнским организмом через плацентарное кровообращение, способ получения питательных веществ и кислорода усложняется. Он определяется градиентом диффузии и величиной площади плацентарной мембраны. Артериальная кровь зародыша характеризуется признаками физиологической' гипоксе-мии, степень ее насыщения кислородом не превышает
50—60%. Для плода она является нормальной и не вы зывает состояния тканевой гипоксии. Физиологическая гипоксемня через хеморецспторы синокаротидной зонг рефлекторно вызывает генерализованные двигательные реакции разгибательного характера. Таким образом некоторое ограничение величины площади плядентар ной поверхности лимитирует поступление питательны} веществ и кислорода из материнской крови в феталь нуго. Двигательные реакции эмбриона ускоряют ско рость кровообращения плаценты.
Различные препятствия в дыхательных путях (глот ке, трахее, бронхах) нарушают дыхание, в результат чего наблюдается респираторный стресс. Сниженш уровня кислорода в воздухе (в горах, стратосфере) вы зывает горную болезнь. Однако главным фактором еле дует считать уменьшение количества кислорода в клет ках (гипоксия) и в крови (гипоксемня). Иногда эя явления сочетаются с накоплением углекислого газа i клетках, поэтому проявляющийся синдром слраведлив< называют гиноксическим гиперкапническим.
Л. В. Серова с соавт. (1973) поднимали крыс в вен тилируемой барокамере на высоту 10 000 м. В первое опыте выжило 6%, после второго подъема — 51'%. Дс гидратация снижала процент смертности. Эти исследо вания подтверждают работы П. Д. Горизоптова (1967)' Barton (1968). Авторы отмечали увеличение концентра ции гемоглобина, эритроцитов и снижение их осмотиче ской резистентности. Гипоксяческне пробы нндуциро вали стрессовые реакции с изменениями функции над почечников. Bertach et al. (1973) вызвали у крыс ги поксическис-гиперкапцнческие расстройства при содер Жаиии их в замкнутых камерах с постепенно изменяю; щимся газовым составом. Гистохимически наблюдал! уменьшение содержания аскорбиновой кислоты в на,а почечниках, нейросекрета в нейрогипофизе. В аденоги-пофизе количество эозинофилов было снижено, в т< же время наблюдали гипертрофию базофилов и их ядер В других опытах крыс подвергали одновременно охлаж дению и гипоксии.
В гипоксическом стрессе наблюдаются различны! изменения фушдаи желез внутренней секреции, от ко торых зависят резистентность и приспособление к кпе лородной недостаточности.
Показано, что после гипофизэктомии животные тяжело переносят гипоксию. Значение имеют не только адренокортнкотропиый, но и тиреотропиый и сомато-тропный гормоны. Тиреоидные гормоны и катехолами-ны участвуют в энергетическом обмене организма и потреблении кислорода, и тем самым играют значительную Роль в устойчивости животных к гипоксии. Кастрированные крысы более резистентны к кислородному голоданию, чем интактные.
Однако следует отметить, что выраженная гипоксия вызывает функциональные изменения в семенниках, снижение сперматогенеза и функции яичников. В то же время анаболические гормоны (кортикостероиды, половые гормоны, инсулин) активизируют синтез белка и восстанавливают запасы жирных кислот, увеличивают содержание неэстерифицированных жирных кислот, повышают резистентность к гипоксемии.
A. В- Захаров (1973) показал, что работа операторов, не требующая физических усилий, сопровождается увеличением легочной вентиляции до 13 л в минуту, ростом потребления кислорода до 150%, снижением парциального давления углекислого газа в альвеолярном воздухе при одновременном снижении потребления кислорода.
Согласно А. В. Волкову (1973), повышение резистентности самок и кастрированных самцов к острой гипоксии объясняется влиянием эстрогенов, снижающих функцию щитовидной железы и активизирующих ре-гпкулоэндотелиальную систему. В процессах дезинтоксикации одновременно наблюдали повышение функции коры надпочечников и секреторной деятельности панкреаса.
B. С. Лифшиц (1949), В. Н. Зворыкин (1951), М- П. Бресткин (1952), Н. Н. Сиротинин (1954) отмерли нарушения поведенческих и условно-рефлекторных реакций при нарастающей кислородной недостаточно-:ти. Ван Лир и соавт. (1967) наблюдали, что при ги-
оксическом стрессе у людей возникают отрицательные моции.
Л. В. Калюжный и Г. П. Гороян (1973) обнаружили,
по при гипоксическом стрессе реакции самораздраже-
я згнетались, облегчалась реакция избегания. С по-
Щью электрофнзиологического метода авторы обна-
ружили возбуждение в ааднемедиальных отделах гипс таламуса, адренергических формациях и ядр^ подкорки.
Таким образом, при гипоксическом стрессе нараст ет выделение тиреоидных гормонов, а затем корти* стероидов.
6.5.2. Гипероксический, гипербарический стресс
В настоящее время накоплен опыт о влиянии гип ■барических камер на организм человека. В них на(Я дается повышение концентрации кислорода в кро что благоприятно действует при тяжелых операш (на сердце, дыхательном аппарате и др.).
Экспериментально интересно сочетать гипероксик| повышением атмосферного давления или создать, дель кессонной болезни. С другой стороны, при под! ных стрессовых явлениях наблюдаются также расстр ства функций надпочечников.
Гипербарическая оксигеннзация, как пока! Б. В. Петровский с соавт. (1977—1978), несмотря собственный стрессорный характер, повышает pel тентность организма при сложных операциях, предст ляющих собой сильные стрессорные воздействия, пербарическая оксигеннзация применяется в последИ годы в клинической и экспериментальной хирурги! большим успехом при лечении тромбозов и эмболии судов различных органов и головного мозга (Е. И. зов с соавт., 1978).
Однако введение кислорода рассматривают как! тистрессовый фактор, усиливающий вторую фазу стр«
6.6. ФИЗИЧЕСКИЕ СТРЕССЫ
Выше мы говорили об изменениях атмосферного1 ления как стрессорном факторе. К ним можно лриба! космические излучения, магнитные поля, явление не! мости и др. В настоящее время накоплены интерес данные о влиянии магнитных полей на различные ф цнн организма. Выявлены изменения функции еердй сосудистой и вегетативной нервной систем, состава к и др.
Dinculescu (1963), Simagi, Bernathini (1966) и др. в начальном периоде действия магнитного поля обнаружили повышение активности коры надпочечников, однако при длительных воздействиях (до 35 суток) наступала атрофия пучковой зоны. Е. А. Савин с соавт. (1973) исследовали действие постоянных магнитных полей на белых крысах и подтвердили данные цитированных выше авторов. Одновременно в лимфоидных органах, в тимусе и селезенке они наблюдали уменьшение количества фолликулов и лимфоцитов. Особенно большие изменения возникали после 15-суточного воздействия магнитного поля.
В опытах М. С. Кахана с соавт. (1965) под влиянием магнитного поля у собак отмечали изменения в биоэлектрической активности ядер передней и задней частей гипоталамуса. В некоторых случаях явно повышалась активность адренергических формаций и щитовидной железы.
Известно, что антигравитационные рефлексы в космосе исчезают, нарушается функция вестибулярного аппарата, поэтому при невесомости наступает вестибулярный стресс. Экспериментально подобный стресс можно вызвать у животных разрушением лабиринтов, введением в полость лабиринтов ватки, смоченной 10'%-ным раствором новокаина. 3. С. Донцова, И. И. Григоренко (1973) наблюдали через 30 мин хаотичность в моносинаптических ответах, расстройство экстензорных и флексорных рефлексов, реципрокной иннервации. Как видно, некоторые физические воздействия, такие как магнитные волны, влияют на гипога-ламические центры.
6.7. КОСМИЧЕСКИЙ СТРЕСС
Космический стресс состоит из разных компонентов. В начале, наблюдается перегрузка, а в дальнейшем в зависимости от длительности полета и тренировки космонавта — влияние следующих факторов: эмоций, невесомости, гиподинамии, вестибулярных нарушений.
Наиболее целенаправленные исследования по космической физиологии осуществлены В. В. Париным (1968), О. Г. Газенко (1974) и др. Они показали возможность прогнозирования в космической биологии. В
дальнейшем Р. М. Баевский (1978—1980) уточнил при* ципы исследования адаптации организма к условия* длительного космического полета. Им установлен прог ностический аспект понятий «здоровье», «болезнь» «адаптация». Показано, что степень напряжения perjj ляторных механизмов — один из фундаментальных дикаторов текущего состояния организма. Автс считает, что состояние кровообращения является показг телем адаптационной деятельности организма. Соглас но. Р. М. Баевскому, прогнозирование ортостатическо| устойчивости организма — ценный элемент космическс медицины. Можно прогнозировать переносимость стибулярных воздействий, степень умственного напр* жения и утомления в космических полетах. В. Н. Mad ников, А. А. Лосев, О. П. Козеренко, Л. Г. Мордовска^ (1973) показали, что воздействие перегрузок в течен! 6 и 8 дней по 21 мин ежедневно вызывает эмоционал^ ные поведенческие и вегетативные реакции, изменен!; пульса, дыхания и кожно-гальванического рефлекс При поперечно направленных перегрузках у летчик^ наблюдаются психофизиологические стрессовые реа| ции. С помощью чехословацкого прибора «Оксимет^ космонавты исследовали кислородный режим ткане мозга.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|