- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 14 страница
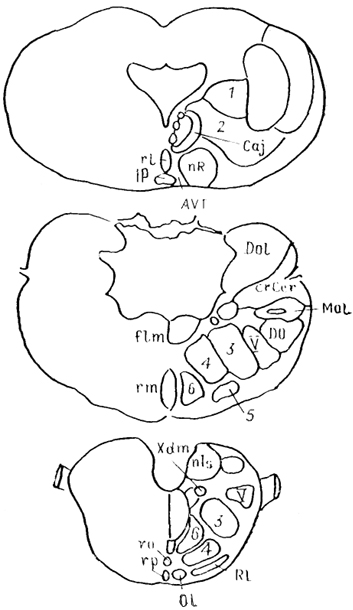
Рис. 62. Ядра ретикулярной формации ската Platyrhinoidis triseriata (Stuesse е. а., 1990).
1 - клиноподобное ядро, 2 - nucl. subcuneiformis, 3 - мелкоклеточное, 4 - гигантоклеточное, 5 - парагигантоклеточное н 6 - крупноклеточное ретикулярные ядра.
Отметим, что для обобщающих заключений о роли медианной зоны ретикулярной фюрмации в ряду позвоночных данных пока явно недостаточно, так как экспериментальное изучение этого отдела выполнено на небольшом числе представителей, а физиологические эксперименты вообще фрагментарны.
* Медиальная часть некоторыми авторами описана как мелкоклеточное ядро шва, nucl,. raphes parvocellularis, или ядро шва, nucl. del raphe.
* У птиц на уровне среднего мозга иногда выделяют линеарное ядро, nucl. linearis.
§ 2. МЕДИАЛЬНАЯ ЗОНА РЕТИКУЛЯРНОЙ ФОРМАЦИИ
Медиальная зона ретикулярной формации наиболее обширна и составляет непрерывный тяж клеток среднего и большого диаметра, занимающих обширные области базальной пластинки. В целом ее описывают как часть тегментума ствола мозга, не включенную в какие-либо ядерные комплексы краниальных нервов или в известные релейные центры. В составе медиальной зоны обнаруживается специфический тип клеток - так называемые ретикулярные нейроны. Они обладают определенным характером дендритных ветвлений и аксонами, которые за
счет бифуркации образуют две противоположно направленные ветви, отдающие по пути многочисленные коллатерали. Такие аксоны образуют значительное число синапсов, активация которых обеспечивает одновременность воздействия клетки на множество других.
У хрящевых рыб медиальная зона занимает всю протяженность ствола и образована крупными нейронами, лежащими вентролатерально по отношению к МОП. В ее составе многими авторами описан ряд ядер (хотя следует отметить, что у некоторых видов, например Scyliorhinus canicula, выделить ядра вообще не удалось, поскольку эта часть ретикулярной формации представлена клеточной зоной равномерной плотности). Так, в мозге некоторых акул и скатов различают три самостоятельных клеточных скопления в ромбовидном мозге и одно - в пределах среднего; последнее во многих работах названо ядром медиального продольного пучка (рис. 60). .
Ромбенцефалический отдел медиальной зоны содержит нижнее, среднее и верхнее ретикулярные ядра, nucll. reticularis inferior, medius et superior. Основываясь на результатах холологических исследований и анализе распределения дендритов нейронов, можно полагать, что афференты этой области образованы структурами спинного мозга, тектума, ядрами мозжечка. В отношении эфферентов, экспериментально не изученных, существует предположение (на основании соотношения количества волокон МПП и числа крупных нейронов ретикулярной формации, лежащих рострально от уровня отсчета), что большой объем проекций адресуется спинному мозгу в составе МПП, система которого, как и вся медиальная ретикулярная формация, прекрасно развита у пластиножаберных. Ядро МПП составлено диффузно лежащими группами клеток и расположено рострально от ядра III нерва. Все ядра медиальной зоны у акул и скатов содержат относительно крупные 3-4-полярные нейроны. Наиболее крупные нейроны, уступающие по размерам лишь маутнеровским клеткам, образуют медиальное ядро. Поступление многих афферентных влияний в эту часть ретикулярной формации обеспечивается дендритами самих ретикулярных нейронов, проникающими в пределы соседних структур или проводящих путей. Тем не менее у одного из видов Ginglymostotna обнаружен прямой теленцефалический вход в область, которую гомологизируют с ретикулярным ядром . перешейка других позвоночных, но которую в силу ее слабой оформленности описывают под другим названием - "ядро Н".
В мозге лучеперых рыб в пределах медиальной зоны выделяют две совокупности клеток - ромбенцефалическую и расположенную в ростральной части ствола область, которая некоторыми авторами описана как ядро МПП. Первая располагается вентролатерально по отношению к МПП и латерально ограничена ядрами бранхиальной моторной колонны. В ее состав
входят три агрегации: нижнее, среднее и верхнее ретикулярные ядра.* Нижнее ретикулярное ядро, nucl. reticularis inferior, соседствует с моторными ядрами IX и X пар черепномозговых нервов. Среднее ретикулярное ядро, nucl. reticularis medius, лежит на уровне вхождения корешка VIII нерва; его каудальная часть расположена медиально от моторного ядра VII нерва, а ростральная - медиально от моторного ядра V нерва. В пределах среднего ретикулярного ядра выделяют промежуточную, пре- и постмаутнеровскую части, однако выраженность каждой из них варьирует, и не во всех случаях они четко представлены. Верхнее ретикулярное ядро, nucl. reticularis superior, лежит в ростральной области ромбенцефалического отдела ростро-медиально по отношению к моторному ядру V нерва.
В целом медиальная часть ретикулярной формации лучеперых рыб содержит большое число крупных нейронов. В мозге Ameiurus обнаружено 20 крупных нейронов, расположенных вблизи октаво-латеральной области и подобных мюллеровским клеткам миног; 14 пар гигантских нейронов описаны в соответствующей области мозга у Anguilla. Эти клетки составляют один из основных (в мозге низших) нисходящих цереброспинальных путей.
При исследовании афферентов медиальной зоны обнаружено, что длинные, ориентированные на периферию дендриты ее нейронов формируют синаптические контакты с волокнами, восходящими из спинного мозга, а также с эфферентами тектума. У костистых рыб, кроме того, бульбарная часть ретикулярной формации получает проекции от сенсорных ядер V, VII, IX и X черепномозговых нервов (преимущественно контрлатеральной стороны). Для среднего ретикулярного ядра основными являются афференты от вестибулярного комплекса. В медиальную ретикулярную зону приходят также коллатерали эфферентов октаво-латеральной области и многие волокна церебелло-тегментальной системы. Эфференты ромбенцефалического отдела входят в состав ипси- и контрлатерального медиального продольного пучка и направляются к моторным ядрам черепномозговых нервов (в частности, V и VII) и к спинному мозгу. Часть эфферентов формирует восходящие проекции ретикулярной формации, которые, как правило, образованы теми же элементами, что и нисходящие, благодаря бифуркации аксонов.
Мезенцефалическая часть медиальной зоны представлена ядром медиального продольного пучка. В этой же области у некоторых рыб располагаются мюллеровские клетки. Сведения об афферентах этого отдела немногочисленны, но показаны входы от тектума и тела мозжечка. Эфференты проходят в составе МПП.
В составе медиальной зоны ретикулярной формации у амфибий также выделяют нижнее, среднее и верхнее ретикулярные ядра * (в мозге хвостатых амфибий последнее, по-видимому, отсутствует). В области перешейка у амфибий к этой зоне относят также nucl. reticularis isthmi, которое у аксолотля не включают в состав ретикулярной формации, а в области среднего мозга - ядро медиального продольного пучка. От всех ядер данной зоны берут начало нисходящие в спинной мозг проекции (описаны у Rana, Bufo и Xenopus). О восходящих проекциях сведения отсутствуют.
Медиальная зона ретикулярной формации рептилий на основании цитоархитектонических данных описывается разными авторами различно. Тем не менее, в ряде классификаций сделаны попытки гомологизировать ядра этой зоны с имеющимися у птиц и млекопитающих. В пределах ромбовидного мозга рептилий, как и у низших позвоночных, выделяют нижнее, среднее и верхнее ретикулярные ядра,** в области перешейка - ретикулярное ядро перешейка, рострально от него лежит ретикулярная формация среднего мозга, относительно которой многое остается неясным.
Нижнее ядро рептилий разделяют на вентральную и дорсальную части. Вентральная часть отсутствует у черепах, в простой форме она обнаружена у крокодилов и некоторых ящериц, но наиболее развитой оказывается у тейид и сцинков и у всех змей. Полагают, что эта часть ядра в филогенезе появляется в мозге рептилий относительно поздно - после отхождения черепах от основного ствола, т. е. у текодонтов, и в наибольшей степени развивается у тех змей и ящериц, которые обладают хорошо выраженной аксиальной мускулатурой. Считается также, что эквивалент данной части присутствует и в мозге млекопитающих. Среди небольшого числа известных афферентов наиболее значительными являются входы ипсилатерального тектума, точное место их окончания не определено. Исходя из аналогий с известными функциями гомологичной части ретикулярной формации млекопитающих полагают, что она вовлечена в контроль аксиальной мускулатуры, а отсюда становятся понятными ее слабая выраженность у ящериц с недостаточно развитой осевой мускулатурой (агамы, игуаны), отсутствие у черепах и сильное развитие у тех чешуйчатых, которые обладают развитой осевой мускулатурой. Дорсальная часть нижнего ретикулярного ядра получает входы от акустической области, а также
(за счет коллатералей текто-бульбарного тракта и спинального лемниска) от спинного мозга и контрлатерального тектума. Эфференты прослеживаются до люмбо-сакральных отделов спин-лого мозга.
Тесно связано с нижним ядром вентролатеральное ретикулярное ядро, nucl. reticularis ventrolateralis, обнаруженное только у змей и тейид. Во многих случаях его присутствие коррелирует с развитием вентральной части нижнего ядра. Однако у сцинков и гекконов при хорошо выраженной последней вентролатеральное ядро отсутствует. Полагают, что оно могло возникнуть независимо в разных группах рептилий.
К среднему ретикулярному ядру приходят афференты от многих образований: акустических и вестибулярных ядер, спинного мозга, ипси- и контрлатерального тектума, торуса, вентрального стриатума. Дендриты нейронов данного ядра, внедряясь в проводящие пути, получают информацию от волокон спинального лемниска, церебелло-тегментального тракта, МПП. Эфференты ядра адресованы спинному мозгу и проходят в вентральных канатиках. В качестве его эквивалента рассматривается каудальное мостовое ядро млекопитающих, стимуляция которого приводит к активации мышц туловища и конечностей.
Верхнее ретикулярное ядро содержит латеральную и медиальную части. Латеральная часть получает сенсорные входы от латеральной петли, спинного мозга, тектума, торуса, а также афференты вентрального стриатума. Ее Эфференты направляются по дорсолатеральным канатикам в спинной мозг. Медиальная часть верхнего ретикулярного ядра изучена менее детально. Известны ее афференты от тектума, идущие в составе предорсального пучка, а также Эфференты, адресованные шейным отделам спинного мозга. Во многих отношениях это ядро сходно с клиноподобным ретикулярным ядром среднего мозга млекопитающих.
Ядра медиальной зоны ретикулярной формации у птиц и млекопитающих включают множество образований. В продолговатом мозге располагаются гигантоклеточное и парамедианное ядра, nucll. reticularis gygantocellularis et paramedianus, рострально от них лежат оральное и каудальное ретикулярные ядра моста, nucll. reticularis pontis oralis et caudalis, а также ретикулярное ядро покрышки моста, nucl. reticularis tegmenti pontis, которое у птиц носит название nucl. papillioformis,* а на уровне среднего мозга обширная область ретикулярной формации образует клиноподобное ядро, nucl. cuneiformis, и дорсальное тегментальное ядро Гуддена, nucl. tegmenti dorsalis (Gudden).
Афферентами ретикулярной формации ромбовидного мозга
являются коллатерали восходящих сенсорных путей, а также волокна спинно-ретикулярного тракта. У млекопитающих, кроме того, все образования получают коллатерали волокон пирамидного тракта, а также проекции ядер мозжечка и гипоталамуса, волокна которых идут в составе пучка Гуддена. Афференты мезенцефалического отдела также представлены сенсорными входами, однако в отличие от нижележащих уровней, здесь возрастает число афферентов стрио-паллидарного происхождения.
Особенностью связей ретикулярного ядра покрышки млекопитающих и его гомолога у птиц являются выраженные входы от преоптической области и эфференты, адресованные мозжечку, - структуры, по-видимому, связанные с осуществлением зрительно-кинетических нистагмов, в частности - горизонтальных.
Эфференты медиальной зоны ретикулярной формации этих животных, подобно другим позвоночным, образуют восходящие и нисходящие проекции. Последние формируют ретикуло-спинальные связи, а также связи с моторными ядрами черепномозговых нервов. Пути, направленные в спинной мозг, происходят из многих ядер. Так, это основные проекции клиноподобного ядра, нейроны которого оказывают влияние главным образом на шейные сегменты спинного мозга. Это ядро часто описывают как "среднемозговую локомоторную область" за его способность вызывать шагательные движения при стимуляции у децеребрированной кошки. Вместе с тем многие ретикулярные нейроны являются одновременно источниками и восходящих проекций (в частности, нейроны гигантоклеточного ядра).
Восходящие проекции ретикулярной формации мозга млекопитающих привлекли внимание специалистов в конце 1940-х гг., когда была обнаружена способность ретикулярной формации оказывать влияние на характер ЭЭГ. Последовавшие за этим морфологические исследования показали, что аксоны крупных нейронов медиальной части ретикулярной формации (гигантоклеточное поле) проецируются рострально в структуры конечного мозга. Сформированное в результате этих работ представление о ретикулярной активирующей системе (РАС) в дальнейшем было многократно подтверждено физиологическими экспериментами. С точки зрения морфологов представления о структуре и компонентах РАС также были расширены (в частности, сюда была включена система ядер шва), однако подробные исследования локализации терминалей ретикулярных нейронов единичны.
Показано (на крысах), что восходящие проекции поступают к промежуточному и конечному мозгу в составе центрального тегментального тракта, медиального продольного пучка и пучка Фореля. Восходящие проекции орального и каудального ядер моста оказываются во многих отношениях сходными: они оканчиваются в поле Фореля и неопределенной зоне, дорсальном таламусе и субталамических образованиях, а также в областях, где имеет место зрительно-моторная конвергенция, - претектальная область, промежуточные отделы передних холмов, ядра глазодвигательного комплекса,
вентральная часть латерального коленчатого тела. Последняя группа структур является основной мишенью проекций орального ядра моста. Другая его особенность состоит в выраженности связей с лимбическими структурами - интерпедункулярным ядром и мамиллярными телами, а также с компактной частью черной субстанции. Мезенцефалическая часть восходящих проекций, направляется к неспецифическим ядрам талагуса, передним холмам, латеральному коленчатому телу, ЦСВ среднего мозга и субталамусу. Имеются также сведения о том, что по крайней мере 10-15% восходящих волокон, адресованы непосредственно коре.
Наконец, среди ядер медиальной зоны выделяется группа, которая в качестве единственной мишени своих эфферентов имеет мозжечок. К ней относятся латеральное, парамедианное и ретикулярное тегментальное ядра покрышки моста.
Таким образом, медиальная зона ретикулярной формации мозга млекопитающих - это важнейший источник нисходящих проекций, осуществляющий контроль сегментарных моторных структур. Другой системой связей, опосредуемой в основном этим отделом ретикулярной формации, является ретикулярная активирующая система.
Нейрохимические методы позволили выявить черты сходства между разными позвоночными, дополнительные к тем, которые рослежены на высших позвоночных по классическим морфологическим критериям. Так, анализ распределения моноаминергических нейронов показал, что многие их группы, в том числе локализованные в пределах медиальной зоны ретикулярной формации, оказываются сходными у самых разных позвоночных, в. частности голубое пятно и вентральная тегментальная область.
Голубое пятно млекопитающих, locus coeruleus, - это небольшое, лежащее в перивентрикулярной области на уровне моста ядро, которое имеет пигментированные клетки и составлено в основном норадренергическими нейронами (группа А6моноаминергических нейронов). Система его проекций чрезвычайно широка: каждый нейрон за счет сильного ветвления аксона может контактировать с множеством образований. Однако в пределах ядра, по-видимому, существует некая приуроченность проекций. Так, значительная часть нейронов голубого пятна помимо норадреналина содержит один или два нейропептида. Согласно существующим данным такие нейроны проецируются не на кору и спинной мозг, а на гипоталамус. Другие же клетки образуют связи, восходящие к слоям I-II новой коры, гиппо-кампу, амигдале, перегородке, стриатуму и коре мозжечка. Нисходящие эфференты адресуются спинному мозгу, где формируют густые сплетения вокруг мотонейронов, а также терминируют в области симпатических ядер и в дорсальном роге.*
Сравнительно недавно аналогичное образование было выявлено в мозге хрящевых рыб при исследовании топографии и
нейрохимических характеристик. У многих лучеперых рыб в области покрышки перешейка также описана группа катехоламинергических клеток, которые посылают проекции в молекулярный слой коры мозжечка и расцениваются некоторыми отдельными авторами как гомолог голубого пятна (у мормирид оно описано под названием "ядро Q"). У рептилий и птиц голубое пятно, насколько известно, имеет такие же, как у млекопитающих, топографию, медиаторную характеристику и общие черты организации связей.
Другой областью, включенной в пределы медиальной зоны ретикулярной формации млекопитающих, является вентральная тегментальная область, area ventralis tegmenti. Она представлена несколькими небольшими группами главным образом до-фаминергических клеток, лежащих вблизи средней линии в основании среднего мозга.* Ее границы цитоархитектонически устанавливаются приблизительно: дорсолатерально от нее располагается красное ядро, латерально-черная субстанция, а вентро-каудально - интерпедункулярное ядро. В числе источников афферентации дорсальное ядро шва, голубое пятно, ретикулярные ядра моста, черная субстанция, ядра мозжечка, ядра поводков, гипоталамус и лимбические отделы теленцефалона.
В составе вентральной тегментальной области находится группа А10 дофаминергических клеток, которая вместе с компактной частью черной субстанции рассматривается как исходная структура мезокортикальной системы. Эфференты AVT классифицируются на пять групп: мезо-ромбенцефалические связи, распределяемые среди ядер ствола и мозжечка; мезо-диенцефалические пути, адресованные таламусу и гипоталамусу (особенно медиодорсальному ядру таламуса и передней гипоталамической и преоптической областям); проходящие вместе мезо-стриатарные и мезо-лимбические проекции, адресованные вентральному стриатуму, перегородке, обонятельному бугорку, некоторым отделам амигдалы, гиппокампу; мезо-кортикальные пути, адресованные различным областям новой коры.**
Полагают, что в мозге млекопитающих вентральная тегментальная область является одним из важнейших звеньев, обеспечивающих взаимодействие лимбической и моторных систем и таким образом участвующих в организации системных поведенческих реакций.
Вентральная тегментальная область была описана и в мозге рептилий и птиц и идентифицирована на основании как медиаторных
характеристик (дофаминергические нейроны), так и сходства связей, в частности адресованных конечному мозгу. У низших позвоночных она описана в качестве перипедункулярной области, area peripeduncularis, содержащей у амфибий и рыб как дофамин- так и серотонинергические нейроны. У некоторых костистых рыб и рептилий обнаружены обширные восходящие проекции в паллиальные области конечного мозга.
* Ранее часто описывались как области моторного ядра покрышки, nucl. Ttegmenti motorius.
* Ретикулярная формация амфибий детально не изучена, и к ним обычно применяют одну из многочисленных номенклатур, разработанных для рептилий.
** Вся ромбенцефалическая часть ретикулярной формации рептилий, согласно разным классификациям, описывалась в качестве единой структуры - как моторное ядро тегментума, или как колонна В. Возможно, что отсутствие четких границ и явилось причиной объединения всех названных ядер в одно образование.
* Отметим, что многие ядра ретикулярной формации птиц получили названия, подобные таковым у млекопитающих, без доказательства их гомологии.
* Основные афференты приходят в голубое пятно от парагигантокле-точного ядра, предлежащего ядра XII нерва и ЦСВ среднего мозга.
* В состав вентральной тегментальной области входят следующие ядра: nucll. paranigralis, interfascicularis, parabrachialis pigmentosus, linearis cauda-lis et linearis rostralis.
** Эта группа эфферентов значительно различается у млекопитающих разных групп: у хищных и приматов обнаруживается максимальная приуроченность входов к префронтальной, орбитофронтальной и цингулярной коре; у грызунов описано диффузное распределение проекций.
§ 3. ЛАТЕРАЛЬНАЯ ЗОНА РЕТИКУЛЯРНОЙ ФОРМАЦИИ
Латеральная зона ретикулярной формации состоит из мелких нейронов, которые у низших образуют колонну вблизи пограничной борозды, и не имеет четко выраженных границ. Лишь в области расположения ядра солитарного тракта и моторного ядра тройничного нерва они составляют медиальную и латеральную границы этого отдела ретикулярной формации.
У хрящевых рыб латеральная зона представлена среднего размера би- и триполярными нейронами. У некоторых акул в ее пределах у места вхождения VIII нерва описано "ядро В", от которого берут начало волокна, осуществляющие иннервацию рецепторного отдела октаво-латеральной системы. У скатов Raja также описано "ядро В", оно лежит рострально от моторного ядра тройничного нерва. Афференты к нему, возможно, поступают от мозжечка в результате ветвления дендритов ретикулярных нейронов среди волокон церебелло-бульбарного тракта. Эфференты направлены к спинному мозгу.*
У актиноптеригий соответствующая область составлена из мелких и средних нейронов. Полагали, что этот отдел, названный латеральной частью ретикулярного ядра, nucl. reticularis pars lateralis, обеспечивает проведение импульсации к эффекторным отделам мозга от вкусовой и соматической систем.
Далеко не у всех изученных видов рептилий латеральная зона разделена на ядра. Так, у каймана в области ромбовидного мозга выделены две клеточные группы - L1и L2Среди черепах Chelonia разделение не проводится, тем не менее у Chrysemis описаны ретикулярное латеральное ядро, nucl. reticularis lateralis, вероятный гомолог L1 , и латеральная часть моторного ядра покрышки, nucl. motorius tegmenti pars lateralis, которое гомологизируют с L2 каймана. Однако цитоархитектонических критериев явно недостаточно для разделения этой части ретикулярной формации на ядра и для последующего их сопоставления с имеющимися у других животных. Так, выделенное лишь на основании нейронных особенностей латеральное ретикулярное
ядро, как показано в экспериментальных исследованиях, получает у черепах восходящие спинальные афференты, что позволяет соотнести его с латеральным ретикулярным ядром, или ядром боковых канатиков, млекопитающих.
Следует отметить, что в более поздних работах отвергается адекватность подобного разделения применительно к черепахам: ядро, описанное как латеральное ретикулярное, у данного вида не выражено, а латеральная часть моторного ядра покрышки является каудальным полюсом моторного ядра VII нерва. Другие авторы на основании сопоставления характера связей и нейронных особенностей относят к латеральной зоне рептилий вентролатеральное ретикулярное ядро, nucl. reticularis ventrolateralis, описанное в мозге тейид и змей и отсутствующее у других ящериц, черепах и крокодилов (рис. 63, а). Возможно, что такое разнообразие мнений, отличающее именно оценку латерального отдела ретикулярной формации, объясняется в первую" очередь отсутствием четких критериев его выделения, связанным с невыраженностью топографических границ.
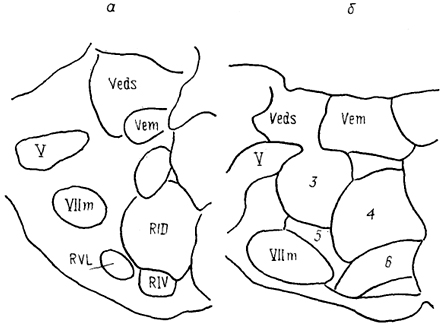
Рис. 63. Ядра ростральной части продолговатого мозга ящерицы Tupinambis nigropunctatum (а) и крысы Rattus norvegicus (б) (Cruce, 1984).
RID - дорсальная и RIV - вентральная части нижнего ретикулярного ядра, RVL - вентролатеральное ретикулярное ядро. 3-6 - см. рис. 62.
У птиц и млекопитающих к латеральной зоне относят участки, сосредоточенные вблизи сенсорных ядер, а также обширные скопления ретикулярных нейронов в пределах сенсорных образований. Наиболее крупным скоплением этой зоны в продолговатом мозге является латеральное ретикулярное ядро, nucl. reticularis lateralis, или ядро латерального канатика, nucl. funiculi lateralis. Оно присутствует у всех животных этой группы и у многих состоит из главной и субтригеминальной частей. У некоторых же видов (в частности, летучая мышь нетопырь) - это единственное четко выраженное ретикулярное образование. Его
афференты подходят со всех уровней спинного мозга, а основной поток эфферентов адресован мозжечку. Другими ядрами этой зоны являются мелкоклеточное ретикулярное ядро, nucl. reticularis parvocellularis (у птиц оно описано как сплетение Хорсли, plexus Horsley), и центральное ядро продолговатого мозга, nucl. medullae oblongata centralis (рис. 61). На уровне моста их сменяет латеральное ретикулярное ядро моста, nucl. reticularis pontis lateralis.* Общими афферентами к этим ядрам являются проекции спинного мозга. Другие поступают от соседних с ретикулярными сенсорных ядер. На уровне среднего мозга к латеральной зоне относят парабрахиальные и педункуло-понтийное ядра. Эфферентные нейроны этой зоны в основном дают начало коротким путям, направленным в медиальную ретикулярную формацию (на этом основании часто латеральную ретикулярную формацию рассматривают как релейное звено в сенсорно-моторных взаимодействиях). Некоторые ядра (парабрахиальные, педункуло-понтийное), причисляемые к этой зоне, вероятно, входят в состав мозговых систем и приурочены прежде всего к лимбической и экстрапирамидной системам.
В целом принято рассматривать латеральную зону ретикулярной формации как наиболее молодую ее часть, которая опосредует, по крайней мере у млекопитающих, взаимодействия многочисленных сенсорных образований с моторными системами мозга. Нечеткость информации о специфических признаках этой зоны не позволяет создать полное представление как о ее составе и функциях у млекопитающих, так и о ее наличии и степени развития у других позвоночных. Возможно, что те ретикуло-подобные элементы, которые в мозге млекопитающих обнаруживаются в ретикулярном ядре таламуса, латеральной и задней областях гипоталамуса, в некоторых слоях новой коры, также могут рассматриваться как отдел ретикулярной формации, однако для подобного вывода необходимы четкие, в том числе и функциональные, критерии.
* Относительно круглоротых сведения отсутствуют, хотя возможно, что описанное в их мозге ядро Янсена - место конвергенции входов соматической и октаво-латеральной систем - также правомерно отнести к числу ретикулярных.
* Возможно, к этой области могут быть отнесены предлежащее ядро XII нерва и ядро Колликера-Фузе.
§ 4. СРАВНИТЕЛЬНЫЙ АНАЛИЗ РЕТИКУЛЯРНОЙ ФОРМАЦИИ ПОЗВОНОЧНЫХ
На основании сравнительно-морфологических исследований высших позвоночных некоторые авторы делают вывод о существовании так называемых "базовых" ядер ретикулярной формации, присутствующих у всех представителей данного класса или нескольких классов. При этом высказывается гипотеза о возрастании их числа в ходе эволюции, хотя признается, что некоторые ядра независимо утрачиваются. В качестве обоснований приводятся данные об общем увеличении числа ядер, в частности "базовых", у млекопитающих по сравнению с рептилиями.
Такая точка зрения противоречит ранее высказанному мнению о постепенном упрощении ретикулярных отделов в связи с прогрессивным развитием высших отделов мозга. Результаты исследований последних лет показали, что ретикулярная формация низших позвоночных действительно оказывается четко структурированной (данные на хрящевых рыбах с использованием нейрохимических методов) и в ней обнаруживаются многие из тех групп, которые, как считалось ранее, характерны лишь для млекопитающих (рис. 62).
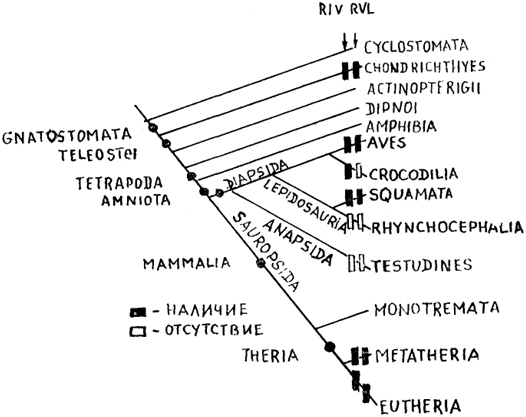
Рис. 64. Представленность вентральной части нижнего (RIV) и вентролатерального (RVL) ретикулярных ядер в мозге позвоночных (Cruce, Newman, 1984).
Однако, у этих же представителей низших позвоночных отсутствуют те отделы ретикулярной формации, которые преимущественно включены в многокомпонентные системы мозга или приурочены к его филогенетически молодым отделам (например, nucl. raphes dorsalis).
В другом подходе, позволившем провести анализ ретикулярных образований разных позвоночных, учитывался комплекс признаков, включая детальную организацию их нисходящих проекций (рис. 64). Действительно, согласно традиционным взглядам, крупные нейроны ствола мозга позвоночных формируют проекции к моторным отделам мозга, в том числе спинного, и составляют таким образом уровень, координирующий деятельность моторных ядер. Многочисленные исследования подтвердили эту точку зрения, и в результате ретикуло-моторные
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|