- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 13 страница
Черная субстанция, substantia nigra, - характерная для мозга млекопитающих структура, лежащая в вентральной части покрышки. Она разделяется на компактную и ретикулярную части, partes compacta et reticulata, соответственно. Компактная часть у человека и высших приматов имеет специфическую пигментацию, обусловленную наличием дофаминергических нейронов (у других млекопитающих менее выражена). Тем не менее локализация ядра у всех млекопитающих сходна: вентральная часть среднего мозга у низших млекопитающих, не имеющих компактных ножек мозга, и положение между тегментумом и ножками мозга у видов, где последние хорошо развиты. Каудально черная субстанция простирается до мостовых ядер. Связи двух отделов черной субстанции различны. Ретикулярная часть получает входы от стриатума и проецируется в тектум, таламус и педункуло-понтийное ядро. Компактная часть имеет множество источников афферентации, среди которых миндалевидный комплекс, ядра шва, хабенулярные ядра. Эфференты адресованы стриатуму. Таким образом, именно ретикулярная часть черной субстанции может рассматриваться в качестве аналога вышеупомянутых ядер, опосредующих стрио-тектальные связи. В целом же черная субстанция расценивается как одно из важнейших звеньев экстрапирамидной системы млекопитающих.
Гомологом черной субстанции у птиц, рептилий и бесхвостых амфибий считают покрышечное педункуло-понтийное ядро, nucl. tegmenti pedunculopontinus, лежащее в вентролатеральной области среднемозгового тегментума; у некоторых рептилий (например, ящерицы) его называют, как и у млекопитающих, черной субстанцией. Связи этого ядра характеризуются двусторонними проекциями к стриатуму и таламусу, а также зфферентами, адресованными тектуму и ретикулярной формации ствола. Помимо сходства связей имеется и сходство-нейрохимических характеристик: в составе данного ядра обнаружены катехоламинергические нейроны. При этом у птиц, они сосредоточены в компактной части педункуло-понтийного' ядра, которую считают гомологом одноименного отдела черной; субстанции, гомологом же ретикулярной части у них является дорсальное покрышечное педункуло-понтийное ядро.
Сведения относительно гомологичных черной субстанции структур в мозге других позвоночных фрагментарны. Так, у акул в ростральной части среднего мозга находится скопление-мелких нейронов - латеральное ядро покрышки, nucl. tegmenti lateralis.* Известно, что оно имеет двусторонние связи с тектумом и афференты от спинного мозга. Некоторые авторы рассматривают латеральное ядро покрышки как примордиум черной субстанции.
Покрышечное педункуло-понтийное ядро описано и в мозге млекопитающих, однако по характеру связей оно отличается от одноименного ядра других позвоночных тем, что адресованные ему афференты происходят из вентральной части стриатума и ретикулярной части черной субстанции. Кроме того, характерными являются двусторонние связи с субталамическим ядром. Не вызывает сомнений принадлежность покрышечного педункуло-понтийного ядра к экстрапирамидной системе млекопитающих. Некоторые авторы в качестве предшественника этого отдела рассматривают интерколликулярное ядро рептилий, которое характеризуется входами от вентральной части стриатума и эфферентами к ретикулярной формации и тектуму.
Таким образом, часть структур среднего мозга, расположенных в области покрышки, по-видимому, является звеньями многокомпонентных систем мозга, в частности моторных. У млекопитающих эти структуры наиболее подробно изучены, о других животных сведения в основном фрагментарны. Недостаточность представлений о системной организации затрудняет гомологизацию входящих в эти структуры образований. Отметим, впрочем, что эта организация может значительно отличаться: если у млекопитающих важнейшим премоторным центром, передающим влияния теленцефальных отделов на спинной мозг, является крупноклеточная часть красного ядра,
то у других, особенно у высших позвоночных, эта роль может принадлежать глубоким слоям тектума, и таким образом сами, системы окажутся состоящими из разных компонентов - ситуация, в которой, вероятно, установление гомологии нельзя считать правомочным.
На уровне среднего мозга имеется структура, свойственная мозгу всех ныне живущих позвоночных, гомологичность которой не вызывает сомнений, хотя наши представления относительно ее роли далеко не полны. Речь идет об межножковом, или интерпедункулярном ядре, nucl. interpeduncularis. Оно хорошо выражено уже в мозге круглоротых и хрящевых рыб, где располагается в области шва на границе перехода перешейка в средний мозг. Описания этого ядра у пластиножаберных противоречивы, поскольку не все авторы его разграничивают от ядер шва. У высших позвоночных интерпедункулярное ядро латерально ограничено ножками мозга. У всех позвоночных ядро образовано мелкими нейронами с небольшим объемом перинуклеарной цитоплазмы. В пределах структуры размеры нейронов варьируют, что является одним из оснований для выделения нескольких клеточных групп: так, у кошек выделяют 4 или 5, а у крыс от 4 до 7. У миног и акул деление структуры производят по топографии: ромбенцефалическая и мезенцефалическая части у миног, дорсальная и вентральная - у акул. Медиаторные характеристики нейронов ядра также служат основанием для разделения его структуры.
В мозге всех позвоночных основной вход к интерпедункулярному ядру образуют нейроны поводковых (хабенулярных) ядер эпиталамуса. Их аксоны формируют центральную часть отогнутого пучка Мейнерта, fasc. retroflexus Meynert, и, сложным образом перекрещиваясь, входят в межножковое ядро. В исследованных случаях обнаружен сложный и упорядоченный характер организации хабенуло-интерпедункулярных связей. Так, в интерпедункулярном ядре бесхвостых амфибий афференты пронизывают определенным образом ориентированные дендриты нейронов, что приводит к активации практически всей структуры в ответ на стимуляцию одной части хабенуло-интерпедункулярного тракта. Четкая топологическая закономерность обнаружена и в организации связей медиального ядра поводков и интерпедункулярного ядра у млекопитающих.
Другие источники входов в интерпедункулярное ядро известны далеко не в полной мере. Для некоторых животных обнаружены входы от тектума. У млекопитающих среди источников афферентации ядро диагональной полоски Брока, дорсальное ядро покрышки, мезенцефалическое ядро шва, центральное серое вещество и голубое пятно.
Эфференты интерпедункулярного ядра у двоякодышащих рыб и хвостатых амфибий образуют интерпедункуло-бульбарный тракт, отсутствующий у лучеперых рыб. У анура эфференты
ядра направляются к дорсальному и глубокому ядрам покрышки. У млекопитающих, изученных в этом отношении более детально, они адресованы дорсальному ядру покрышки, медиодорсальному ядру таламуса, септальным ядрам, переднему ядру мамиллярных тел, ядру диагональной полоски Брока, преоптической области, вентральному ядру покрышки и латеральному ядру поводков. У некоторых видов обнаружены лроекции в гипоталамус и к центральному ядру шва.
Характерной особенностью нейронов данной структуры является значительное число нейросекреторных элементов. Они обнаружены у некоторых видов, относящихся к разным классам, и, возможно, эта черта свойственна всем позвоночным. Так, у лягушек нейросекреторные клетки содержат крупные гранулы, а отростки имеют крупные варикозные расширения. Дендриты, ветвящиеся в плоскости, перпендикулярной пучку Мейнерта, образуют по всей длине щелевые контакты и заканчиваются на субпиальной поверхности, контактируя с межножковой цистерной. Подобные клетки описаны и в мозге человека, где их отростки формируют концевые ножки на сосудах и пиальной оболочке. Наличие нейросекреторных элементов, а также сильная васкуляризация ядра (по-видимому, свойственная всем позвоночным, имеющим систему кровоснабжения мозга), позволяют объяснить системный характер влияний этой структуры на различные формы поведенческих реакций. Действительно, сначала только на основании анализа связей, а позднее - экспериментальных исследований, был сделан вывод о принадлежности межножкового ядра к структурам лимбической системы, которые опосредуют передачу импульсации на премоторные центры - дорсальное и глубокое тегментальные ядра. Показано участие этих отделов в обеспечении избегания, полового и (у высших) эмоционального поведения. Раннее созревание этого ядра в онтогенезе позволило предположить, что ряд поведенческих реакций новорожденных млекопитающих (например, сосание и глотание) также опосредуется активацией этого ядра. Полагают, что одним из способов осуществления влияний интерпедункулярного ядра на другие структуры мозга может быть выброс биологически активных веществ (в частности, соматостатина) в церебро-спинальную жидкость. Отметим, что довольно сложной оказывается и нейрохимическая характеристика интерпедункулярного ядра; здесь обнаружены нейроны и терминали различной медиаторной природы. Высказывается предположение, что в ряду позвоночных, особенно млекопитающих, происходит нейрохимическая специализация ядра, что может отражать общее прогрессивное развитие лимбической системы у высших позвоночных. Однако недостаточность сведений не позволяет окончательно сделать такой вывод, поскольку у некоторых видов в пределах ядра обнаружены медиаторы, отсутствующие у других, что, вероятно, свидетельствует о существовании специфических путей и характеристик системы.
В общем, интерпедункулярное ядро у всех позвоночных характеризуется целым рядом сходных черт: связями с ядрами поводков, гипоталамусом, дорсальным тегментальным ядром, сильной васкуляризацией; наличием нейросекреторных элементов, граничащих с цереброспинальной жидкостью, которые свидетельствуют о его принадлежности к системам мозга, ответственным за организацию целостных поведенческих актов (у высших - к лимбической системе).
Отметим, что упомянутое выше дорсальное тегментальное ядро, nucl. tegmenti dorsalis, или ядро Гуддена, у всех наземных позвоночных характеризуется наличием афферентов от интерпедункулярного
ядра и в свою очередь проецируется на моторные ядра краниальных нервов, что позволяет рассматривать его как одно из "выходных" звеньев лимбической системы, обеспечивающих доступ к исполнительным отделам мозга.*
Другой структурой, свойственной мозгу большинства позвоночных, за исключением акул** и круглоротых, является ядро перешейка, nucl. isthmi. Наиболее характерная особенность этой структуры - двусторонние связи с тектумом, которые имеют топологически организованные входы и определенные закономерности становления в онтогенезе. У амфибий, некоторых рептилий и млекопитающих наблюдается сочетанное присутствие в тектуме прямого ипсилатерального входа сетчатки и билатеральных проекций ядра перешейка. В то же время отсутствие ипсилатерального ретинального входа совпадает с отсутствием билатеральных проекций. Показано также, что появление активности нейронов ядра перешейка в ходе эмбрионального развития (у амфибий - в период, предшествующий метаморфозу) приурочено к появлению ипсилатеральных сетчаточных проекций. Данное обстоятельство трудно объяснить однозначно, поскольку волокна упомянутых трактов адресованы разным отделам тектума: ипсилатеральные входы - отделам, находящимся в глубине тектума, а вход ядра перешейка - отделам, лежащим на поверхности тектума, над контрлатеральными ретинальными волокнами. Однако если один из этих компонентов отсутствует, то отсутствует и другой. Такая ситуация характерна для большинства птиц и лучеперых рыб, лишенных ипсилатеральных сетчаточных входов и контрлатеральных проекций ядра перешейка на тектум.
У рептилий ядро перешейка дифференцируют на крупно- и мелкоклеточную части. Первая наиболее развита у крокодилов, ящериц, черепах. У змей, лишенных, по мнению большинства авторов, ядра перешейка, в ростральной части покрышки среднего мозга выделяют nucl. tegmenti posterolateralis, которое на основании сходства связей рассматривают как гомолог (аналог?) ядра перешейка (некоторые авторы описывают ядро перешейка в его обычной локализации).
У птиц различают ядро перешейка, nucl. isthmi, и оптическое ядро перешейка, nucl. isthmoopticus. Первое делят на мелко- и крупноклеточные компоненты и полулунное ядро, раrtes parvo- et magnocellularis et nucl. semilunaris. Для всех компонентов этого комплекса показана конвергенция входов зрительной (от тектума) и слуховой (от ядер латеральной петли)
систем. Доказано его участие в реакции сужения зрачка на свет (например, в ответ на звук). Оптическое ядро перешейка описано не у всех видов (например, у Ibis оно замещено полулунным ядром, его дорсальным продолжением). Оно располагается в дорсолатеральной части покрышки и, как обнаружено, принимает участие в центрифугальном контроле чувствительности контрлатеральной сетчатки.
У млекопитающих гомологом ядра перешейка считают-nucl. parabigeminalis, его же рассматривают как гомолог мелкоклеточной части ядра перешейка рептилий и полулунного^ ядра птиц. Это ядро представляет собой небольшую группу клеток, занимающую латеральное положение в среднем мозге, вентрально от медиального коленчатого тела и ручек задних холмов. В ряде случаев в его составе выделяют несколько клеточных групп. Общей чертой всех исследованных в этом отношении видов являются двусторонние связи этого ядра с той частью передних холмов, которая получает ретинальные входы. При этом, если тектальные афференты преимущественно ипсилатеральны, то для обратных связей характерна билатеральность. Учитывая существующую ретинотопию, ряд авторов рассматривает ядро parabigeminalis как сателлитное для передних холмов. Таким образом, nucl. parabigeminalis, подобно ядру перешейка всех позвоночных, оказывается тесно связанным с мезенцефалическим зрительным центром (тектум или передние холмы) и обеспечивает поступление в него информации от контрлатеральной одноименной структуры. Однако, если у большинства позвоночных таким способом, вероятно, обеспечиваются бинокулярные взаимодействия, то у млекопитающих, которые имеют ипсилатеральные ретинальные проекции* возможны и другие функции ядра. Отметим, что у одного вида галаго обнаружены проекции nucl. parabigeminalis в латеральное коленчатое тело, что может быть свойственно и другим животным. Еще одна возможная роль этого ядра - усиление разрешающей способности зрительной системы через активацию истмо-ретинальных проекций.*
Другой структурой, описанной в среднем мозге позвоночных (за исключением круглоротых), является интерколликулярное ядро, nucl. intercollicularis, лежащее в центральной области тегментума. Оно получает спинальные входы и проецируется на таламус. У рептилий оно часто рассматривается как место конвергенции акустических и соматических входов. Иногда его включают в состав торуса в качестве перитораль-ной области, regio peritoralis. Вместе с тем, у некоторых животных ядро с таким названием имеет совершенно другие связи.
Так, у птиц одноименная структура рассматривается как часть тектума, и ее связи в основном адресованы нижележащим структурам. У акул и скатов интерколликулярное ядро располагается в каудальной части покрышки среднего мозга; оно получает билатеральные входы от тектума и проецируется на спинной мозг, являясь, таким образом, релейной структурой для текто-спинальных связей. Наконец, отметим, что у рептилий название "интерколликулярное ядро" дано двум разным структурам, одна из них, вероятно, опосредует связи стриатума с тектумом.
У млекопитающих выделяют как самостоятельное образовавание центральное серое вещество (ЦСВ) среднего мозга, substantia grisea centralis mesencephali, которое, как полагают, гомологично перивентрикулярной области тектума других позвоночных. Однако связи этих образований значительно различаются. ЦСВ млекопитающих чрезвычайно гетерогенно, содержит несколько типов нейронов, характеризуется разнообразием медиаторов. В его составе выделяют ряд отделов, отличающихся в том числе и по связям. Оно получает многочисленные входы, источники которых локализованы во всех отделах ЦИС. Значительная часть этих структур относится к лимбической системе (в том числе медиальная префронтальная кора, амигдала, гипоталамус, латеральное хабенулярное ядро), а также к звеньям моторных систем (моторная область коры, мозжечок, черная субстанция). Наконец, на уровень ЦСВ проецируются сенсорные структуры: ядро солитарного тракта, главное ядро тройничного нерва, зрительные и слуховые центры среднего мозга, нейроны пластин IV и V спинного мозга, Эфференты адресованы многочисленным структурам, в числе которых лимбические отделы (латеральный гипоталамус, амигдала, фронтальная область коры), парафасцикулярный комплекс таламуса, ядра шва, парабрахиальная и дорсолатеральная области покрышки моста. Многие из этих связей приурочены к определенным областям ЦСВ, однако высокая степень коллатерализации его эфферентных нейронов, а также отсутствие детальных исследований на разных систематических группах не позволяют представить детальную картину организации связей ЦСВ.
Тем не менее, имеющиеся данные позволяют отнести этот отдел мозга млекопитающих к структурам лимбической системы. Так, показано, что его тесные связи с гипоталамусом обеспечивают протекание некоторых фаз репродуктивного поведения. Существует мнение, что ЦСВ является структурой такого уровня, где происходит конвергенция лимбической и моторной систем мозга. Значительное число фактов свидетельствует о причастности ЦСВ к системе эндогенной анальгезии. Это же подтверждают характер поступающей информации от ноцицептивных элементов спинного мозга, наличие опиоидных рецепторов,
а также результаты экспериментального изучения влияния ЦСВ на восприятие боли. Однако значительная медиаторная гетерогенность не позволяет пока создать детальную схему взаимодействий этого отдела с другими структурами ноцицептивной системы. Наконец, к настоящему времени накопилось множество данных о его участии в организации видоспецифических вокализаций, которые, как полагают, являются акустическим выражением эмоционального состояния. Это :может вполне согласовываться со сведениями о наличии ноцицептивных входов и афферентов от вышележащих лимбических образований. Таким образом, возможно, что ЦСВ является одним из "выходных" образований лимбической системы, которое преимущественно активирует голосовой аппарат животных и регулирует поступление болевой информации в мозг. Такое сочетание данных представляет интерес и в качестве доказательства справедливости представления о том, что вокализация в процессе эволюции возникает как средство выражения внутреннего состояния (возможно, по принципу смещенной активности) и лишь позднее приобретает коммуникационные функции.
Сложный характер связей и неполнота представлений о роли данного отдела затрудняют проведение сравнительно-анатомического или функционального исследования, поскольку речь идет о структуре, включаемой в состав многокомпонентной лимбической системы млекопитающих, сведения о наличии :и уровне развития которой в мозге других позвоночных отсутствуют.
Таким образом, на уровне среднего мозга у всех позвоночных представлены сенсорные образования зрительной и октаво-латеральной систем. Число моторных образований невелико, и они контролируют сокращение экстраокулярных мышц. Многочисленные связи с исполнительными отделами обеспечиваются на этом уровне надсегментарными (премоторными) образованиями: глубокими слоями тектума, ядром МПП, дорсальным сегментальным ядром, красным ядром, на которых происходит конвергенция входов от сенсорной и лимбической систем, определяя тем самым важную, а в ряде случае решающую роль этого отдела в организации системных поведенческих реакций.
* Отметим, что в составе пучка проходят и восходящие проекции, в; частности соединяющие вестибулярный и глазодвигательный комплексы, а также связи между ядрами III, IV и VI черепномозговых нервов.
* Латеральное ядро покрышки у Hydrolagus ранее описано как nucll dorsalis thalami или nucl. rotundus mesencephali.
* У млекопитающих усложнение лимбической системы приводит и к усложнению связей ядра, но их характер остается прежним: афференты - от лимбических структур, эфференты - к моторным и вегетативным ядрам.
** В мозге акул ядро с таким названием описано, но кроме сходства в местоположении других данных, свидетельствующих о гомологичности структуры, не имеется.
* На уровне среднего мозга располагаются и другие структуры, связанные с обработкой зрительной информации. Они входят в состав добавочной зрительной системы и будут, описаны в главе 7, поскольку включают и диенцефальные образования.
ГЛАВА 6
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ
Об эволюционных преобразованиях ретикулярной формации известно еще немного. Сложность такого анализа объясняется, целым рядом обстоятельств. Естественно, что для проведения, сравнительно-анатомического исследования необходимо опираться на единую номенклатуру ретикулярных отделов, что в свою очередь подразумевает наличие исчерпывающих доказательств гомологии описываемых структур. Вместе с тем исследования ретикулярной формации до недавних пор ограничивались в основном классическими морфологическими подходами - описанием топографии, топологии, положения в мозге относительно постоянных анатомических образований, морфологии нейронов. Таким образом, суждения о гомологии отдельных: образований зачастую базируются лишь на некоторых морфологических чертах сходства.
Вопрос о трансформации ретикулярных образований в эволюции сложен еще и в силу неразработанности критериев для отнесения структур к числу ретикулярных. Действительно в ретикулярную формацию в мозге низших включают отделы, которые не относятся к системе ядер черепномозговых нервов. В мозге высших помимо сенсорных и моторных ядер появляются: многочисленные надсегментарные образования (вышележащие структуры сенсорных систем, премоторные отделы и релейные центры), которые также не считают принадлежащими ретикулярной формации. Однако, согласно имеющимся сведениям, многие из них имеют ретикулярное происхождение. К настоящему времени появились исследования нейрохимических: характеристик ретикулярных образований у представителей разных таксонов позвоночных, что позволило существенным образом расширить представления о преобразованиях ретикулярной формации ходе эволюционного развития, но такие исследования пока немногочисленны.
В современной литературе ретикулярную формацию ствола амниот обычно подразделяют на три продольные зоны: медианную, медиальную и латеральную (исходя из имеющихся к
настоящему времени данных, подобное разделение оказывается справедливым и для анамний). Медианная зона состоит из клеток, расположенных по средней линии тегментума ствола мозга и составляющих область шва, а также лежащих вблизи нее; медиальная наиболее значительна по объему у всех позвоночных и составляет срединную часть ствола мозга; латеральная в основном приурочена к области сосредоточения сенсорных ядер, а у низших располагается вблизи пограничной борозды.
§ 1. МЕДИАННАЯ ЗОНА РЕТИКУЛЯРНОЙ ФОРМАЦИИ
Медианная зона ретикулярной формации состоит из рассеянных нейронов, формирующих более или менее четко выраженные скопления (см. рис. 26, 60). Так, уже в мозге круглоротых в этой области выделяются переднее, среднее и заднее ядра шва, nucll. raphes anterior, medius, posterior. Медианная зона акул представлена мелкими клетками, которые в ростральной части образуют скопление, гомологичное верхнему ядру шва, nucl. raphes superior, других позвоночных (рис. 34). В каудальной части медианной зоны клетки лежат диффузно и не составляют ядра. У некоторых представителей хрящевых рыб (скатов Raja) в каудальной части обнаруживается достаточно четко выраженная агрегация нейронов больших размеров - крупноклеточное нижнее ядро шва, nucl. raphes inferior magnocellularis. Оно получает тектальные входы и проецируется на спинной мозг, а у окатов иннервирует электрические органы. У костистых рыб также выделяют ядра шва (см. рис. 35, а), при этом имеются данные (y Lepomis) об их серотонинергической природе. У Xenopus в составе медианной зоны выделяют
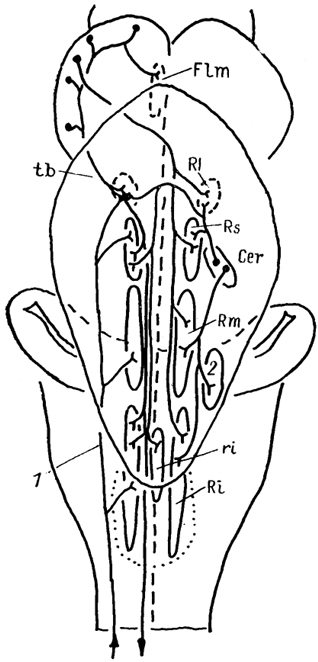
Рис. 60. Афференты ретикулярной формации ствола мозга хрящевых ствола мозга хрящевых рыб (Smeets e. а., 1983).
1 - спинномозговые афференты, 2 - ядро В.
единое ядро шва с двумя частями - вентральной и дорсальной. Первая, nucl. raphes pars ventralis, состоящая из мелких нейронов, простирается от obex до перешейка вентрально или в пределах внутренних дугообразных волокон. Дорсальная часть ядра, nucl. raphes pars dorsalis, проходит выше .дугообразных волокон и имеет большую протяженность: от каудальной части интерпедункулярного ядра рострально и ниже obex - каудально. Имеются данные о проекции этих ядер на спинной мозг.
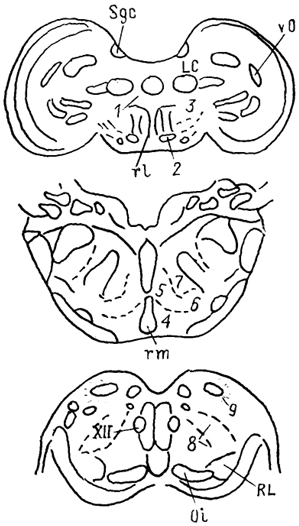
Рис. 61. Ретикулярная формация птиц (по: Pearson, 1976).
1 - nucl. annularis, 2 - nucl. popillioformis, 3 - латеральная зона ретикулярной формации, 4 - парамедианное и 5 - гигантоклеточное ретикулярные ядра, 6 - латеральное парагигантоклеточное ядро, 7 - каудальное ретикулярное ядро моста, 8 - центральное ядро продолговатого мозга, 9 - сплетение Хорсли.
Ядра шва у рептилий весьма вариабельны даже у представителей разных семейств одного отряда. Естественно, что чрезвычайно разнообразной оказывается и номенклатура этих ретикулярных образований. Так, в мозге черепах в пределах медианной зоны выделяют нижнее и верхнее ядра шва. Нижнее ядро шва, nucl. raphes inferior, проходит от уровня ядра VI нерва до спинного мозга, его ростральный и каудальный полюсы составлены мелкими нейронами, в средней же части встречается (значительное число крупных клеток, которые не всегда могут быть отграничены от смежного и расположенного сбоку нижнего ретикулярного ядра. Рострально от этого ядра располагается группа крупных клеток. Она обнаружена у представителей некоторых групп рептилий (Chelofiia, Chrysemys) и носит название верхней части крупноклеточного ядра шва, nucl. raphes magnocellularis pars superior. Верхнее ядро шва, nucl. raphes superior, разделено на латеральную и медиальную части.* Оно составлено компактной группой клеток среднего размера и вытянуто в вертикальной плоскости. Латеральная его часть граничит с промежуточным отделом
медиальной части и содержит мелкие нейроны, плотность которых несколько меньше, чем в медиальной части. Ядра шва рептилий получают коллатерали волокон предорсального пучка, возникающего от контрлатерального тектума и проходящего вблизи средней линии. Эфференты области шва спускаются по латеральным канатикам в спинной мозг и доходят до его нижних сегментов. Возможно, что данная область, ретикулярной формации рептилий оказывает регулирующее влияние на спинной мозг, подобно обнаруженному у млекопитающих. Кроме того, показано окончание аксонов в таламусе и гипоталамусе, а также в среднем мозге.
У птиц и млекопитающих в составе медианной зоны ретикулярной формации в пределах ромбовидного мозга выделяют темное, бледное и большое ядра шва, nucll. raphes obscurus, pallidus, magnus, соответственно. Рострально располагается мостовое ядро шва, nucl. raphes pontis, а на уровне среднего мозга - верхнее центральное и дорсальное ядра шва, nucll. raphes superior centralis et dorsalis * (рис. 61).
Афферентами ядер шва являются коллатерали восходящих сенсорных проекций. Эфференты образуют восходящие и нисходящие пути. Последние в основном происходят от большого ядра шва. Восходящие проекции берут начало от всех ядер (но в меньшей степени от большого и мостового ядер) и адресуются структурам .конечного мозга и лимбическим отделам промежуточного и среднего мозга: гиппокампу, амигдале, ядрам диагональной полоски, хабенулярному латеральному ядру, медиальной преоптической области, перегородке, вентральной тегментальной области среднего мозга. От мостового ядра шва начинаются пути в мозжечок.
Экспериментальные исследования показали, что стимуляция ядер шва: оказывает системное влияние на состояние коры большого мозга и опосредует половое поведение, сон, моторную активность, а также протекание таких поведенческих актов, как агрессия, реакция избегания и т. п. Объяснением столь широкой сферы влияний этого отдела ретикулярной формации является, по-видимому, ее принадлежность к серотонинергической системе мозга, в которой наибольший удельный вес принадлежит именно системе ядер шва. Полагают, что серотонинергические терминали могут рассматриваться как экстрасинаптический модулятор, способный контролировать метаболическую активность других систем. Возможно, с этим связаны и специфический характер влияний ядер шва на систему ноцицепции и анальгезирующий эффект их стимуляции. В целом же сложная система связей ядер шва позволяет предположить, что между ними существует разделение по-преимущественному контролю определенных клеточных популяций, и таким образом ядра шва оказываются функционально гетерогенными.
Серотонинергическая природа ядер шва обнаружена к настоящему времени у всех исследованных в этом отношении позвоночных - круглоротых, хрящевых и лучеперых рыб, амфибий и амниот. При этом показано удиви-
тельное сходство многих черт организации связей. Так, уже у круглоротых помимо выраженных нисходящих связей серотонинергические нейроны посылают проекции к стриатуму и паллиуму, к преоптической области, в тектум и дорсальный таламус. Использование нейрохимических методов позволило выделить в мозге хрящевых рыб Platyrhinoidis triseriata большинство ядер системы шва, гомологичных имеющимся у млекопитающих (рис. 62). Исключение составляет дорсальное ядро шва;его отсутствие в мозге хрящевых рыб, возможно, свидетельствует о том, что и эта древняя система мозга преобразовывалась в ходе эволюции: поскольку основной объем связей дорсального ядра шва составляют восходящие в теленцефалон проекции, то, вероятно, развитие и этого звена шло параллельно с формированием новых отделов плаща конечного мозга.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|