- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 12 страница
У актиноптеригий полукруглый валик состоит из ряда слоев, параллельных вентрикулярной поверхности. Его размеры коррелируют с уровнем развития октаво-латеральной системы. При этом каждый из отделов торуса связан с обработкой информации, поступающей в конечном итоге от разных типов рецепторов октаво-латеральной системы. Так, у сома Ictalurus торус состоит из медиально расположенного центрального ядра и латерального ядра; последнее делится на части: латеральную и медиальную - меньшую по размерам. Акустический вход адресован центральному ядру, медиальная часть латерального ядра в основном содержит "электрочувствительные" нейроны, латеральная
же часть является местом проекций механорецепторов органов боковой линии. Еще более сложно дифференцирован торус у мормирид, у которых в его составе выделяют 7 ядер. Таким образом, входы медуллярных образований, относящихся к октаво-латеральной системе, на уровне торуса оказываются четко пространственно расположенными. При этом для каждого из отделов показана соматотопическая организация. Например, ростро-каудальное расположение рецепторов соответствует аналогичному направлению представительства в контрлатеральном торусе у сома Ictalurus. Эфференты торуса у актиноптеригий выяснены недостаточно. У голостей и телеостей они в составе заднего мезенцефало-церебеллярного тракта адресуются телу и; заслонке мозжечка. Комиссуральные связи образуют поперечную комиссуру, com. transversa, лежащую вентрально от промежуточного мозга. Полагают, что часть ее волокон адресована постхиазматической области гипоталамуса.
У бесхвостых амфибий торус представлен крупными парными утолщениями, выступающими в полость среднемозгового желудочка. Они очень велики у Ranidae (см. рис. 30) и смыкаются по средней линии, деля каудальную часть желудочка на дорсальный и вентральный отделы. В составе торуса у разных амфибий выделяют от трех до пяти ядер. У Rana catesbeiana торус включает главное, nucl. principalis, ламинарное, nucl. laminaris, крупноклеточное, nucl. magnocellularis, комиссуральное, nucl. commissuralis, субэпендимное, nucl. subependimalis, ядра. У Xenopus выделены только первые три. Афферентами ядер торуса являются входы от нижележащих образований слуховой системы. Так, главное ядро получает проекции от дорсального октавного ядра и от верхней оливы. Несколько меньший объем входов верхней оливы адресован крупноклеточному и комиссуральному ядрам. Нейроны ламинарного ядра, по-видимому, получают проекции от клеток главного ядра, к которым простираются их дендриты. Помимо входов от слуховых образований в торус амфибий проецируется и комплекс вестибулярных ядер. Кроме того, имеются входы от ретикулярной формации, гипоталамуса, спинного мозга, а также от контрлатерального торуса. Основной объем эфферентов этого отдела направляется к двум группам таламических ядер, а оттуда - в две области теленцефалона. Нисходящие проекции адресованы спинному мозгу. Обнаружена высокая чувствительность нейронов ламинарного ядра к изменению концентрации половых гормонов, что может объяснить участие данного уровня в организации видоспецифических вокализаций в брачный период.
У рептилий существуют значительные вариации в степени развития среднемозговых слуховых центров. У некоторых из; них торус представлен группой клеток, лежащей под тектумом. У змей различают небольшую агрегацию нейронов, рострально переходящую в перивентрикулярную зону тектума. У
других полукруглые валики являются крупными образованиями, внедренными в тектальный желудочек под каудальными 2/3 тектума. Они состоят из субвентрикулярно расположенного ламинарного ядра, nucl. laminaris, и более глубоко лежащего центрального ядра, nucl. Centralis.* Вентролатерально от последнего у некоторых рептилий (например, черепах) выделяют еще один компонент полукруглого валика, который описан под названием интерколликулярного, или периторального отдела, геgio peritoralis, и цитоархитектонически выражен незначительно.
Связи разных отделов торуса оказались несколько различными. Основной вход к центральному ядру поступает от контрлатеральных слуховых отделов ромбовидного мозга в составе латеральной петли. Дорсолатеральная часть, кроме того, получает проекции от цервикальных отделов спинного мозга. Эфференты центрального ядра адресованы ламинарному и таламическим ядрам, связанным с обработкой акустической информации (например, у кайманов - ядра "Z" et reuniens). Основные афференты ламинарного ядра представлены входами от центрального ядра торуса, эфференты главным образом направляются в спинной мозг. На основании характера связей его гомологизируют со среднемозговой ретикулярной формацией млекопитающих. Периторальный отдел является местом перекрытия спинальных и слуховых входов. Источники последних детально не выяснены, но некоторые авторы отрицают наличие афферентов от кохлеарного комплекса. Эфференты этой области адресованы таламическому ядру nucl. reuniens. Периторальный отдел гомологизируют с наружным (латеральным) ядром задних холмов млекопитающих. Особенностью торуса рептилий является существование выраженного входа от вестибулярного комплекса, который поступает вместе с кохлеарными проекциями.
Гомологом полукруглого валика в мозге птиц является дорсальная часть латерального мезендефалического ядра, nucl. lateralis mesencephali pars dorsalis (рис. 58). Ее размеры значительно отличаются у разных видов и достигают максимальной величины у некоторых видов сов и Steatornis.** У многих видов птиц эта структура разделяется на наружное и центральное ядра. Центральное получает входы от нижележащих слуховых образований и в свою очередь проецируется на наружное ядро. Одни эфференты отсюда направляются в таламический отдел слуховой системы, другие же - к тем образованиям, которые связаны с системой вокализации: на уровне среднего мозга птиц - это небольшая группа клеток, сосредоточенных вблизи каудальной части ядер глазодвигательного нерва вентромедиально от желудочка - дорсомедиальное интерколликулярное
ядро, nucl. intercollicularis dorsomedialis. Его эфференты, в свою очередь, адресованы каудальной части ядра подъязычного нерва, осуществляющей иннервацию голосового аппарата.
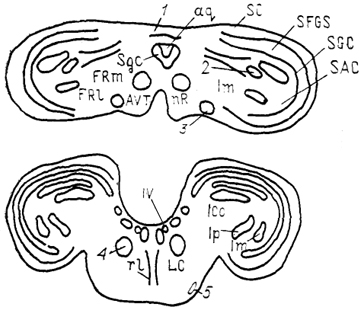
Рис. 58. Строение среднего мозга птиц (по: Miceli, Reperant, 1985).
1 - тектальная комиссура, 2 - дорсальная часть мезенцефалического латерального ядра, 3 - эктомамиллярное ядро, 4 - nucl. annularis, 5 - латеральное ядро моста. Остальные обозначения, как на рис. 52.
Гомологом торуса в мозге млекопитающих являются нижние бугры четверохолмия, colliculi inferiores, или задние холмы, colliculi posteriores. Они составляют каудальную часть крыши среднего мозга, достигая у некоторых видов значительных размеров и превосходя зрительные структуры среднего мозга. У всех представителей класса задние холмы представлены несколькими отделами: центральным ядром, четырех-слойной корой, покрывающей его дорсально и каудально (перицентральное ядро - по другой номенклатуре), комиссуральным ядром и тегментальной пограничной областью, отделяющей задние холмы от передних и образующей дорсомедиальное и латеральное поверхностные ядра Каждый из отделов получает различную комбинацию афферентов от нижележащих слуховых образований: центральному ядру адресован основной объем входов латеральной петли, в коре заднего холма доминируют комиссуральные входы, для тегментальной пограничной области характерно перекрытие акустических и сомато-сенсорных проекций, а также афференты от черной субстанции. Восходящие эфференты составляют ручки задних холмов, brachia colliculorum posteriores, и адресованы таламусу. Связи с центральным серым веществом и ретикулярной формацией обеспечивают участие структуры в организации видоспецифиче-ских вокализаций.
Помимо названных структур на уровне среднего мозга обнаруживается еще одно сенсорное образование - среднемозговое ядро тройничного нерва, nucl. mesencephalicus n. trigemini. Око свойственно всем позвоночным, обладающим челюстями, и представлено группой клеток, лежащей в перивентрикулярной области на всем протяжении тектума и отличающейся от окружающей мозговой ткани размерами и формой нейронов. Развитие структуры варьирует. Так, у мормирид ее можно выделить только по аналогии в расположении и клеточном составе, у других
рыб ядро включает несколько хорошо дифференцированных отделов. Основной сенсорный вход представлен проекциями проприорецепторов челюстных мышц и, в меньшей степени, гактильных рецепторов (у акул, лишенных в области рострума мышечных веретен, вход сформирован только тактильными рецепторами). Клетки ядра образуют большое число плотных контактов с соседними элементами. Кроме того, большинство из них контактирует с цереброспинальной жидкостью полости желудочка, а у акул - и с сосудами области тектума. Другие афференты ядра известны не в полной мере: у акул они сформированы глубокими слоями тектума и, как следует из электрофизи-ологических данных, ядром мозжечка. Эфференты ядра у большинства исследованных групп оказываются подобными. Они направляются к моторному ядру тройничного нерва, в ромбовидный мозг и цервикальные сегменты спинного мозга. Для птиц, млекопитающих и акул, по-видимому, характерны эфференты, адресованные мозжечку.
В некоторых случаях в строении и связях ядра обнаруживаются особенности. Так, у карпа среднемозговоё ядро тройничного нерва не имеет связей с моторными ядрами тройничного и лицевого нервов и, по-видимому, не связано с проприорецепторами мышц области головы. У некоторых амфибий часть входов в это ядро образовано корешком IV черепномозгового нерва. Для пластиножаберных характерны большие размеры клеток и значительный объем дендритных ветвлений. Кроме того, только у данных животных обнаружены контакты клеток ядра с сосудами тектальной области, что может объясняться особенностями кровоснабжения их мозга. Ликворконтактирующие нейроны обнаружены в мозге акул и млекопитающих; аналогичные сведения относительно других позвоночных отсутствуют.
* Показано, что у Lepisosteus в ростро-медиальной части тектума имеется небольшой прямой ипсилатеральный вход.
* В экспериментах показано участие этих трактов в обеспечении таких поведенческих реакций, как обнаружение и захват добычи (за счет поворота головы и последующего укуса - активация перекрещенного и прямого трактов соответственно).
* Соматотопическая карта оказывается так же, как и зрительная, неравномерной: например, область морды у черепах, подобно центральной части сетчатки, представлена на значительно большей поверхности.
* Отметим, что у некоторых змей центральное ядро описано под названием "параторус".
** Интересным примером асимметрии стволовых отделов мозга является преимущественное развитие этого ядра слева у данного вида.
§ 2. МОТОРНЫЕ ЯДРА СРЕДНЕГО МОЗГА
В области тегментума среднего мозга располагается значительное количество образований, относящихся к разным системам мозга. Часть области, примыкающая к тектуму, занята ядрами глазодвигательного (III) и блокового (IV) черепномозговых нервов, nucll. n. oculomotorii et trochlearis соответственно. Вместе с ядром отводящего нерва, nucl. n. abducentis, лежащим в ростральной части ромбовидного мозга (на уровне моста у млекопитающих, в области перешейка у многих других), они образуют единый глазодвигательный комплекс со сложными взаимными связями и едиными функциями. Наружные мышцы глаза, иннервируемые этими ядрами, происходят в эмбриогенезе из разных сегментов, и этим объясняется тот факт, что эти небольшие по размерам мышцы иннервируются разными группами мозговых структур. Сфера иннервации для каждого из ядер оказывается собственной, однако в пределах мозга между
ними устанавливаются тесные двусторонние и четко организованные связи. Нейронный состав ядер достаточно разнообразен и отличает эту группу от ядра подъязычного нерва, также относящегося к группе соматомоторных.
В мозге некоторых круглоротых описаны группы клеток, которые соответствуют глазодвигательному комплексу челюстно-ротых; большой объем связей они получают от вестибулярного комплекса. В мозге хрящевых рыб все ядра глазодвигательного комплекса располагаются около средней линии в непосредственной близости от медиального продольного пучка. При этом ядро VI нерва четко отделено от других. У актиноптеригий оно располагается на ростральном уровне ядра лицевого нерва, и его нейроны иннервируют наружную прямую мышцу глаза. У многих костистых рыб в пределах ядра выделяют две клеточные группы. У амфибий и рептилий ядро отводящего нерва не всегда четко отделено от окружающей ретикулярной формации и в целом сходно с таковым акул и двоякодышащих; для его связей характерно наличие входов от вестибулярного комплекса.*
В составе отводящего нерва у позвоночных (за исключением приматов и человека) описывают добавочное ядро. С использованием современных методов его существование показано у саламандры, ящерицы, лягушки и крысы. Во всех случаях нейронный состав добавочного ядра оказался сходным, его аксоны иннервируют мышцу retractor bulbi. Входы к этому ядру, помимо общих с основным ядром, составляют ядра тройничного нерва, а связи с вестибулярным комплексом практически отсутствуют.
Ядро IV нерва отличается от других ядер комплекса тем, что его эфференты, составляющие блоковый нерв, выходят с дорсальной поверхности мозга. Сферой его иннервации у челюстных является коитрлатеральная верхняя косая мышца, а у бесчелюстных - ее гомолог - задняя косая мышца. Только в мозге круглоротых ядро IV нерва занимает дорсальное положение, у всех остальных оно располагается в вентральном тегментуме. В организации ядра IV нерва у разных позвоночных прослеживается целый ряд общих черт. Так, у всех челюстных дендриты его нейронов входят в область, занятую медиальным продольным пучком. Кроме того, дендриты простираются в область тройничных ядер, ядер октаво-латеральной зоны и таким образом обеспечивают получение влияний от разных отделов и систем мозга. У многих видов млекопитающих, амфибий и у некоторых птиц показано существование и небольшого числа ипсилатеральных проекций, хотя основная масса волокон иннервирует противоположную сторону. Следует отметить, что у млекопитающих
и птиц соотношение числа мышечных волокон и мотонейронов в ядре IV нерва - наименьшее. В связи с этим неожиданными оказались сведения об аналогичном соотношении в мозге миног, что свидетельствует о тонкой и детальной дифференцировке этой системы. Отметим еще одну особенность, касающуюся системы блокового нерва. Речь идет о смешанной природе этого нерва у двух видов хвостатых амфибий - в составе его корешка в мозг входят сенсорные волокна, принадлежащие системе тройничного мезенцефалического нерва и несущие информацию от проприорецепторов, в том числе и глазных мышц. У других животных (бесхвостые амфибии, костистые рыбы, млекопитающие) сенсорные волокна присоединяются к этому корешку экстракраниально, у этих же амфибий они идут вместе с уровня мозга.
Ядро глазодвигательного нерва организовано наиболее сложно и у многих позвоночных состоит из нескольких нейронных групп. Так, в мозге рыб оно разделяется на латеральную и медиальную части. Согласно результатам исследований, проведенных на карпе, ядро обеспечивает ипсилатеральную иннервацию нижней и внутренней прямых мышц,* а также нижней косой мышцы; верхняя прямая мышца получает контрлатеральные входы. У других животных дифференцировка ядра III нерва проведена по-другому,** однако характер распределения сфер иннервации остается сходным. У многих животных дендриты нейронов данного образования, широко ветвясь, устанавливают контакты с областями, занятыми сенсорными ядрами, и таким образом у низших обеспечивается множественность входов. Четко выражены и пути к моторным отделам мозга, в связи с чем становится возможным контроль фиксации глаз при движении тела и головы.
Примером специализации этого отдела являются обнаруженные у некоторых морских телеостей электрические органы, представляющие собой измененные миотомы четырех из шести глазных мышц: медиальной, верхней и латеральной прямых и верхней косой. В этом случае в отличие от других все названные мышцы иннервируются системой III нерва, представительство которого в мозге оказывается гипертрофированным, так как основной его объем составлен нейронами, контролирующими электроциты, и только мелкие клетки осуществляют иннервацию наружных глазных мышц.
Отметим, что в составе III черепномозгового нерва присутствуют и вегетативные волокна, берущие начало от особой группы нейронов, которые формируют у высших парасимпатическое ядро Якубовича, или ядро Эдингера-Вестфаля,- или добавочное ядро III нерва, nucl. accessorius n. oculomotorii. У хрящевых
рыб оно лежит вблизи основного ядра III нерва, но доказательств его парасимпатической природы нет. В то же время у этих животных описан ресничный ганглий и показано участие корешка глазодвигательного нерва в реакции расширения зрачка, у всех других постганглионарные волокна, активируясь, приводят к сужению зрачка.
* У черепах, крокодилов и чешуйчатых имеется дополнительная группа клеток, иннервирующая мигательную перепонку. У гекконов и змей она отсутствует.
* У акул в отличие от других позвоночных внутренняя прямая мышца получает контрлатеральную иннервацию.
** У многих рептилий, млекопитающих и птиц ядро глазодвигательного нерва очень велико и разделяется на дорсальную и вентральную части, состоящие из нескольких отделов.
§ 3. ЯДРА ОБЛАСТИ ПОКРЫШКИ СРЕДНЕГО МОЗГА
Область покрышки среднего мозга и перешейка содержит значительное число образований, далеко не для всех из которых установлены возможные гомологи и выяснены пути преобразования в ходе эволюции. Одна из причин этого - ретикулярное происхождение большинства этих структур: одни тегментальные ядра включены в состав ретикулярной формации, другие, хорошо развитые в мозге высших позвоночных, вероятно, ее производные. Таким образом, становятся понятными многочисленные разночтения и противоречия, имеющиеся в литературе, которая посвящена описанию этих отделов у разных позвоночных.
Тем не менее в составе тегментума перешейка и среднего мозга можно выделить несколько групп образований, сходных в отношении связей и выполняемых функций. Одну из таких групп составляют структуры, обеспечивающие проведение импульсации к моторным ядрам черепномозговых нервов и к спинному мозгу - так называемые премоторные структуры, или надсегментарные моторные отделы. Несмотря на то, что у низших позвоночных эти отделы принадлежат ретикулярной формации, они в отличие от других ретикулярных образований четко представлены в мозге разных позвоночных и могут быть гомологизированы.
В пределах базальной пластинки среднего мозга высших позвоночных располагается крупное скопление клеток - красное ядро, nucl. ruber (рис. 59). Полагают, что данная структура свойственна всем тетраподам, и ее специфической характеристикой являются клетки - источники рубро-спинального тракта, tr. rubrospinalis. Для высших позвоночных идентификация этой структуры не вызывает затруднений. У других же при отсутствии видимых клеточных скоплений (при обработке по Нисслю), тем не менее в области тегментума можно выделить группы клеток со сходными связями. Поэтому в настоящее время для выделения примордиального красного ядра используют несколько критериев: положение в мозге, наличие входов в составе верхних ножек мозжечка и контрлатеральный выход на спинной мозг.
Перекрещенный рубро-спинальный тракт описан у бесхвостых амфибий, четвероногих рептилий, птиц и млекопитающих. Считалось, что у примитивных животных подобные связи отсутствуют. Действительно, у круглоротых не обнаружено тегменто-
спинальных связей, аналогичных рассматриваемым, однако у некоторых рыб в пределах тегментума выделяется группа клеток - источников спинальных связей. Кроме того, у некоторых видов скатов (Raja clavata, Dasyatus sabina) не только четко выделяется группа клеток - примордиум красного ядра, но развиты входы от ядер мозжечка и хорошо выражен перекрещенный рубро-спинальный тракт (заметим, что речь идет о рыбах, интенсивно использующих для движения плавники), о б в
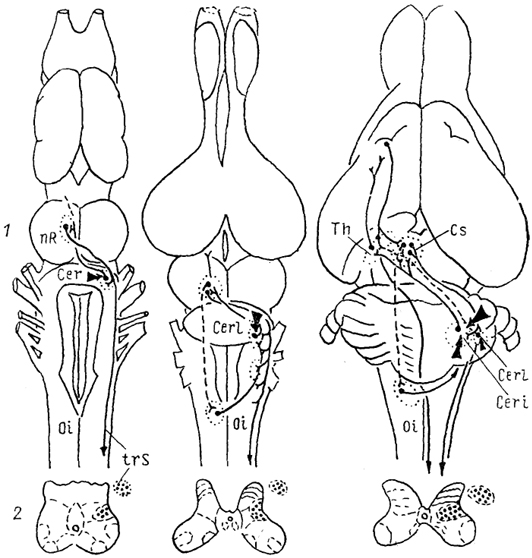
Рис. 59. Эволюция красного ядра (Toth e. а., 1985, Donkelaar, 1988).
а-в - красное ядро бесхвостой амфибии (а), ящерицы (б), опоссума (в). 1 - связи красного ядра, 2 - окончания волокон рубро-спинального тракта.
В мозге бесхвостых амфибий красное ядро выделяется на основе анализа связей: группа клеток, лежащих в вентро-медиальной части тегментума, получает входы от единственного ядра мозжечка через слабо развитые верхние ножки мозжечка и: формирует перекрещенный рубро-спинальный тракт. Пожалуй, бесхвостые амфибии - самые примитивные из групп, обладающих типичным красным ядром. При этом степень развития моторных систем мозга у них далеко не совершенна. Связи с нижней оливой отсутствуют.
У рептилий красное ядро варьирует по размерам. У некоторых
групп оно не может быть цитоархитектонически выделено, у других же хорошо развито, например у четвероногих, и в его составе выделяются клеточные группы, четко отграниченные от окружающей ретикулярной формации. Мозжечковые входы к нему образует латеральное ядро мозжечка. Кроме того, формируются связи от вышележащих отделов - ядер конечного мозга. Эфференты сосредоточены в крупных рубро-бульбарном и рубро-спинальном трактах. Последний проходит через дорсальную часть латерального канатика и оканчивается в латеральном отделе пластин V-VI. В случае отсутствия рубро-спинального сохраняется рубро-бульбарный тракт (например, у питона). Таким образом, у рептилий, особенно имеющих конечности, наблюдается дальнейшее развитие системы красного ядра. Однако связи с нижней оливой на этом уровне еще слабо представлены, равно как и входы от вышележащих уровней мозга.
У птиц в составе красного ядра выделяется по крайней мере два типа нейронов: крупные, сосредоточенные преимущественно дорсомедиально и вентролатерально, и средние (и мелкие), сосредоточенные в основном рострально. Характер связей подобен имеющимся у рептилий.
Только у млекопитающих в пределах данного ядра клетки разных размеров распределены в различных областях структуры, в результате чего в пределах красного ядра описывают крупноклеточную и мелкоклеточную части, partes magno-et parvocellularis. Удельный вес последней и ее связей прогрессивно возрастает, при этом крупноклеточная часть у высших приматов и человека в значительной степени редуцируется. У сумчатых же основной объем красного ядра составлен крупными клетками, а мелкие нейроны еще не сосредоточены в самостоятельную группу. Степень развития красного ядра коррелирует и со способом локомоции: способность к плаванию и полету сочетается с относительно незначительно развитой структурой, а хождение и длинные конечности - с большим по размерам красным ядром и обширными связями его крупноклеточной части. Входы к красному ядру млекопитающих берут начало от разных ядер мозжечка. Уже в мозге примитивных млекопитающих афференты промежуточного, и латерального (зубчатого) ядер мозжечка адресованы разным отделам. Появляются входы от коры мозга. Эфференты также происходят из разных областей: так, у опоссума рубро-спинальный тракт образован крупными клетками, лежащими в каудальной и ростровентральной областях; ростромедиальная и ростродорсальная же образуют рубро-оливарные и рубро-бульбарные проекции соответственно.
У приматов и человека ростральная, наиболее значительная часть, состоящая из относительно мелких клеток, получает входы от новой коры, стриатума и зубчатого ядра мозжечка, а
проецируется на главное ядро нижней оливы и особую группу таламйческих ядер. Крупноклеточная часть имеет незначительное число входов от конечного мозга, основной мозжечковый вход образован нейронами промежуточного ядра. Ее эфференты составляют очень небольшой рубро-спинальный тракт, распределение которого ограничивается верхними сегментами спинного мозга. Очевидно, в ряду млекопитающих происходит постепенное возрастание роли мелкоклеточной части красного ядра, что, вероятно, связано с общим усложнением моторных систем мозга и развитием высших моторных систем, берущих начало с уровня конечного мозга.
Таким образом, крупноклеточная часть красного ядра млекопитающих и в целом красное ядро других позвоночных представляют собой надсегментарный уровень организации моторных систем. Вместе с вестибуло- и ретикуло-спинальным трактами и медиальным продольным . пучком рубро-спинальный тракт обеспечивает организацию многих моторных реакций, в частности локомоции. Развитие же мелкоклеточной части красного ядра, происходящее в связи с усложнением вышележащих отделов, является следствием развития в эволюции высших моторных систем (пирамидной и экстрапирамидной), объединяющих многие отделы мозга.
Отметим, что в мозге круглоротых и всех групп рыб достаточно четко выделяется группа нейронов, образующая моторное ядро покрышки, nucl. motorius tegmenti. Оно является местом конвергенции входов от разных структур, в том числе ют тектума и торуса. Его эфференты проецируются на моторные ядра ствола и спинного мозга. Очевидно, это свойственная низшим премоторная структура аналогична красному ядру; некоторые же авторы рассматривают ее как прекурсор последнего.
Другими структурами, которые можно отнести к премоторным образованиям являются ядро медиального продольного пучка, nucl. fasc. longitudinalis medialis, и промежуточное ядро Кахаля, nucl. interstitialis Cajal. На их нейронах конвергируют входы от разных источников, в том числе от вестибулярного комплекса, тектума (передних холмов), претектальной области. Эфференты формируют часть медиального продольного пучка (МПП), а у некоторых животных - направляются и к ядрам глазодвигательного комплекса. МПП - многокомпонентный путь, свойственный мозгу всех позвоночных. Он соединяет ствол мозга с моторными отделами спинального уровня, характеризуется постоянством расположения и занимает парамедианную позицию в стволе, проходя в вентральных канатиках спинного мозга на большое расстояние. Промежуточное ядро и ядро Кахаля - не единственные, образующие данный путь; на уровне среднего мозга его составляют также эфференты ядра задней комиссуры, а у высших позвоночных и
ядра Даркшевича, и красного ядра. На уровне ромбовидного мозга к нему присоединяются вестибуло-спинальные волокна, а также незначительное число волокон, источниками которых являются сенсорные ядра черепномозговых нервов и мозжечка (показано у рептилий).* Множественность связей определяет участие МПП в таких, реакциях, как сочетанный поворот головы и глаз.
На уровне среднего мозга у разных групп позвоночных имеются и другие структуры, связанные с моторными отделами, однако недостаточность сведений о связях и функциональной роли этих структур не позволяет судить об их гомологичности с топографически сходными ядрами других животных.
Так, у многих позвоночных описано глубокое ядро среднего мозга, nucl. profundus mesencephali, достигающее у некоторых групп значительных размеров (например, у рептилий). Согласно имеющимся данным у акул, амфибий и: рептилий оно получает билатеральный тектальный вход и проецируется на нижележащие моторные структуры. Относительно других позвоночных данные достаточно противоречивы.
В тегментальной области перешейка пластиножаберных описано несколько клеточных групп, обозначенных как ядра F, G, Н. Два первых проецируются на спинной мозг, последнее, возможно, опосредует текто-бульбарные связи, однако недостаточность сведений не дает возможности говорить об их специфичности или гомологии с какими-либо центрами других животных.
Возможно, к этой же группе относится ядро, обнаруженное у всех актиноптеригий и расположенное в вентролатеральной части покрышки. У представителей разных групп лучеперых оно описано под разными названиями: красное ядро покрышки, nucl. ruber tegmenti, латеральное ядро торуса, дорсальная часть энтопедункулярного ядра, nucl. entopeduncularis pars dorsalis. Степень развития ядра варьирует, связи изучены недостаточно, хотя имеются сведения о наличии его эфферентов в составе лобо-бульбарного и лобо-церебеллярного трактов. Отметим, однако, что анализ его связей у голостей позволил предположить, что данная структура является частью торуса.
В числе структур, обеспечивающих связи стволовых отделов; с моторными центрами, у наземных позвоночных выделяются образования, связанные с вокализацией. К ним относится описанное у бесхвостых амфибий nucl. pretrigeminalis (у Хепоpus - дорсальная тегментальная область). Оно располагается в латеральной части тегментума перешейка. Для его нейронов-характерны высокая зависимость от уровня гормонов, выраженные связи с преоптической областью гипоталамуса и наличие
эфферентов, адресованных билатерально ядрам языко-глоточного и блуждающего нервов.
У птиц вентромедиально от желудочка среднего мозга располагается дорсомедиальное интерколликулярное ядро, которое получает афференты от слухового мезенцефалического отдела и адресует эфференты каудальной части ядра подъязычного нерва, осуществляющей иннервацию голосового аппарата.
Помимо премоторных образований на уровне среднего мозга располагаются структуры, выполняющие релейную роль и связывающие между собой различные центры. Такую роль приписывают, в частности, ядру задней комиссуры, nucl. commissurae posterioris, которая у высших позвоночных тесно связана с тектумом. У птиц его вероятным гомологом является ядро, .лежащее в претектальной области, - nucl. spiriformis lateralis thalami. Значительная часть входов к этой структуре у птиц и рептилий образована стриатумом. Таким образом, учитывая особенности контактов, этот отдел рассматривают как звено, опосредующее стрио-тектальные связи, т. е. передающее влияние высших отделов на премоторное образование - глубокие слои тектума, дающие начало нисходящим проекциям. У млекопитающих ядро задней комиссуры обеспечивает связи в пределах ядер глазодвигательного нерва и, как полагают, опосредует движения глаз вверх. Афференты от стриатума не обнаружены, и считается, что аналогичную роль у млекопитающих играет черная субстанция.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|