- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 17 страница
аналогий с делением таламуса других позвоночных. Заключая описание таламуса позвоночных, следует отметить, что в ходе прогрессивной эволюции изменяется степень развития и совершенства разных его отделов: существенно уменьшается область, обозначаемая как синенцефалон, редуцируется область заднего бугорка, а области дорсального (особенно) и вентрального таламуса значительно усложняются, сопровождая преобразования паллиальных отделов мозга.
Поиски аналогов и гомологов в пределах таламуса проводились многократно, однако вряд ли эти попытки можно считать успешными. Достаточно четко у всех челюстных позвоночных в пределах таламуса выделяются лишь ядра, имеющие отношение к проведению сенсорной информации в теленцефалон. Но недостаток, в первую очередь морфологических сведений, а также несовершенство критериев дифференциации не позволяют считать их гомологичными, хотя об аналогах с функциональной точки зрения можно говорить достаточно обоснованно. Заметим, что сложность сопоставления сенсорных ядер обусловлена также и значительным проявлением экологических особенностей (как, например, при делении ЛКТ позвоночных на дорсальный и вентральный отделы). Кроме того, некоторые ядра, традиционно рассматриваемые как сенсорные, имеют выраженные гетеросенсорные характеристики (nucl. reuniens у рептилий), что может расцениваться как свидетельство древних черт организации и, таким образом, ставит под сомнение корректность подобных сравнений.
Еще одной традиционной точкой зрения является утверждение о постеленной редукции вентрального и прогрессивном развитии дорсального таламуса, которое выражается как в усложнении его, так и в возрастании роли последнего. Однако пересмотр разделения таламических образований, проведенный в последние 10-15 лет, привел к "перемещению" ряда отделов из дорсальной части в вентральную и наоборот. Отсутствие общепризнанных критериев разделения таламуса, по-видимому, не дает возможности рассматривать преобразования в таламусе в этом аспекте.
Еще более трудными оказываются поиски аналогий с другими известными ядрами таламуса млекопитающих - ассоциативными и неспецифическими: установить гомологии здесь сложно даже среди млекопитающих, что же касается представителей других классов, то судя по имеющимся данным лишь у рептилий и птиц среди таламических появляются ядра, проецирующиеся на несколько областей паллиума и на этом основании сравнимые с интраламинарными ядрами млекопитающих. У птиц такой структурой является nucl. subrotundus, у рептилий - nucl. perirotundus (у черепах - ядро DM).
Следует отметить, что если в качестве определяющей характеристики таламических отделов принять их связи с теленцефальными образованиями, то прогрессивные изменения в этом отношении в ряду позвоночных очевидны: возрастание объема связей, освобождение их от контрлатеральных путей (имеющихся у хрящевых рыб и бесхвостых амфибий), а также прогрессивное разделение и уточнение теленцефальных мишеней. Аналогичные процессы, вероятно, имеют место и в таламических афферентах. Эти преобразования, по-видимому, происходят и в пределах каждого класса. По крайней мере среди млекопитающих многие из этих особенностей можно проследить на группе неспецифических ядер, которые проходят путь преобразований, параллельный изменениям кортикальных отделов.
Наконец, отметим, что если для таламуса низших позвоночных обычно существование нейронов, проецирующихся на спинной мозг и моторные ядра ствола, то в таламусе высших такие образования в качестве самостоятельных отсутствуют, а структуры,
проецирующиеся на ствол мозга, адресуют свои проекции звеньям надсегментарных или высших моторных систем и, как правило, составляют особые отделы - большинство субталамических образований. Относительно общими остаются для всех позвоночных диенцефальные образования, связанные с организацией зрительно-моторных реакций. Они составляют так называемую претектальную область промежуточного мозга (хотя некоторые ее элементы у низших позвоночных рядом авторов включаются в состав синенцефалона).
Претектальная область располагается на границе промежуточного и среднего мозга. Она содержит многочисленные мелкие группы клеток, образующие ядра претектальной области, nucll. pretectales (Pt). В функциональном и структурном отношениях они оказываются сходными с ядрами добавочной зрительной системы, сосредоточенными в области покрышки среднего мозга. Таким образом, характеристику этих образований, целесообразно проводить параллельно.
Добавочная зрительная система содержит ядра, получающие прямой сетчаточный вход по добавочному (базальному) оптическому тракту. Она представлена у всех челюстноротых несколькими ядрами. Прежде всего это ядро базального оптического тракта, nucl. basalis tr. optici, и зрительное ядро покрышки, nucl. opticus tegmenti (рис. 66, 68). Гомологизация в отношении других образований проведена далеко не полностью.
В мозге рептилий выделяют дорсальную зрительную область и оптическое ядро покрышки среднего мозга, area optica tegmenti mesencephali dorsalis et nucl. opticus tegmenti mesencephali. Последнее было описано под разными названиями: зрительное ядро покрышки, ядро базального оптического тракта, эктомамиллярное ядро.
У птиц на основании различий в нейронном составе выделяют три ядра: дорсальное, латеральное и собственно ядро базального оптического тракта.
У всех млекопитающих в составе добавочной зрительной системы описывают несколько ядер: медиальное, латеральное и дорсальное терминальные, nucll. terminales medialis, lateralis et dorsalis. При этом у примитивных видов максимально развитыми оказываются медиальное терминальное ядро и подходящий к нему нижний пучок добавочной оптической системы (у ехидны это единственный хорошо выраженный компонент системы). У высших наблюдается увеличение других ядер, а у приматов медиальное терминальное ядро и нижний пучок вообще трудно выделить. Полагают, что медиальное терминальное ядро гомологично собственно ядру базального оптического тракта, дорсальное и латеральное терминальные ядра - соответственна дорсальному и латеральному ядрам базального оптического тракта птиц. Этот вывод сделан на основании анализа физиологических характеристик нейронов. По этим же критериям не
которые из ядер оказались весьма близки к структурам претектума, а в отдельных случаях вообще трудно решить вопрос о принадлежности ядра той или иной системе.
В составе претектальной области уже у миног выделяют примордиальное претектальное ядро. У хрящевых и костных рыб здесь описывают несколько ядер, в том числе ядро задней комиссуры.
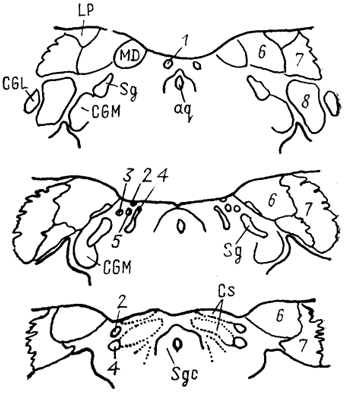
Рис. 76. Претектальная область мозга обезьяны Масаса fascicularis (Kaas, Huerta, 1988).
1-4 - ядра претектальной области: медиальное (1), переднее (2), оливарное (3 и заднее (4); 5 - ядро оптического тракта; 6-8- ядра Pul: медиальное (6), латеральное (7) и нижнее (8).
У высших позвоночных состав претектальной области еще более сложен. На основе цитоархитектоники область подразделяется на несколько групп. Так, у рептилий претектальная область весьма значительно варьирует, в ее составе выделяют медиальную и латеральную группы. Латеральная включает в себя три ядра: коленчатое претектальное, nucl. geniculatus pretectalis, чечевицеобразное среднемозговое, nucl. lentiformis mesencephali и заднедорсальное, nucl. posterodorsalis. Медиальная группа называется чечевицеобразным ядром таламуса, nucl. lentiformis thalami, и разделена на перивентрикулярную (медиальную) и латеральную части, pars plicata et pars extensa. Последняя так же, как ядра латеральной группы, значительно преобразуется у некоторых видов с хорошо развитым зрением (агамы, игуаны, тейиды, вараны, хамелеоны) . У этих животных в составе гипертрофированной pars extensa выделяют дорсальное, вентральное и медиальное претектальные ядра. Последнее, присутствующее также в мозге у черепах и кайманов, описано у них под названием дорсального' ядра задней комиссуры, nucl. dorsalis commissurae posterioris.
Самым крупным ядром претектума птиц является чечевицеобразное мезенцефалическое ядро, nucl. lentiformis mesencephali, в составе которого выделяются подъядра. Кроме того, к этой области относятся диффузное претектальное ядро, nucl, pretectalis diffusus, претектальная оптическая область, area optiса pretectalis и nucl. synencephalicus superficialis.
У большинства млекопитающих в составе претектума выделяют
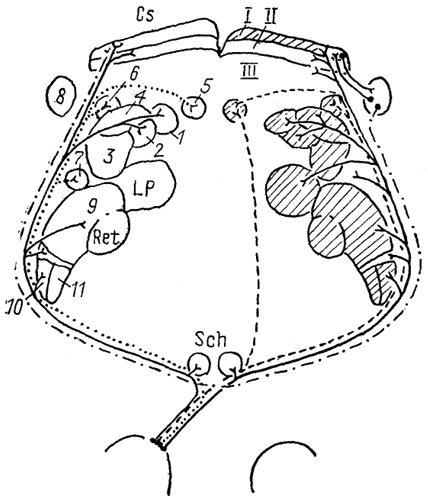
Рис. 77. Схема организации связей структур зрительной системы крысы (Sefton, Dreher, 1985).
1-3 - претектальные ядра: заднее (1), оливарное (2) и переднее (3); 4 - ядро зрительного тракта; 5-7 - терминальные ядра добавочной зрительной системы: медиальное (5), дорсальное (6) и латеральное (7); 8 - nucl. parabigeminalis; 9 - дорсальное ЛКТ. 10-11 - вентральное ЛКТ: крупноклеточная (10) и мелкоклеточная (11) части. I-III - зоны передних холмов: поверхностная (I), зона вхождения зрительного нерва (II), промежуточная (III). Заштрихованы структуры, получающие входы от зрительной области коры.
4-5 ядер. Так, в мозге крыс описывают: ядро оптического тракта, nucl. tracti optici, лежащее среди волокон ручек передних холмов, переднее, заднее и оливарное претектальные ядра, nucll. pretectales anterior, posterior et olivaris (последнее считают гомологом заднедорсального ядра рептилий). Помимо этих ядер в мозге приматов выделяют медиальное претектальное ядро, nucl. pretectalis medialis, нечетко оформленное в мозге грызунов (рис. 76). Полагают, исходя из топографии, хода эмбриогенеза и свойств нейронов, что ядро оптического тракта гомологично чечевицеобразному среднемозговому ядру птиц и рептилий.
У ехидны, вероятно, претектальная область представлена группой клеток, лежащей внутри ручек передних холмов и обозначенной как дорсальное терминальное ядро, или дорсальное терминальное ядро оптического тракта.
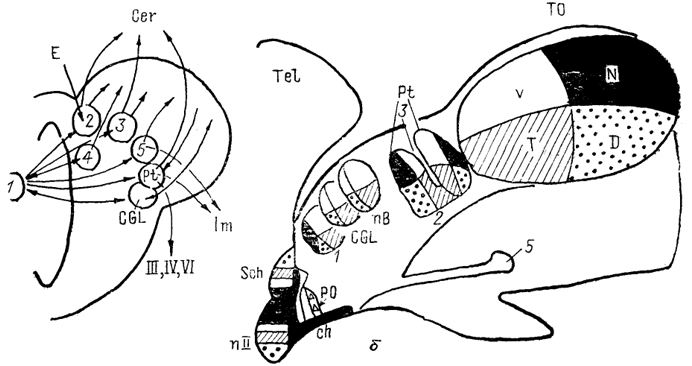
Рис. 78. Схема связей структур, получающих вход сетчатки в мозге костистых рыб (а) и бесхвостых амфибий (б) (по: Vanegas, Ito, 1983; Montgomeri, Fite, 1989).
А: 1 - сетчатка, 2 - ядро задней комиссуры, 3 - претектальная область, 4 - дорсолатеральное ядро, 5 - кортикальное ядро, б: 1 - ростральное зрительное ядро, 2 - заднеталамическое ядро, 3 - крючковидный нейропиль, 4 - лентиформное ядро среднего мозга, 5 - ядро базального зрительного тракта. V, N, Т, D - области представительства в контрлатеральной части мозга вентрального (V), назального (N), темпорального (Т) и дорсального (D) квадрантов сетчатки.
Характер связей структур претектума и добавочной зрительной системы оказался сходным во многих отношениях. Так, пратически все названные ядра получают более или менее выраженный прямой ретинальный вход (для добавочной зрительной системы преимущественно контрлатеральный, для некоторых ядер претектума билатеральный). У птиц и млекопитающих во многие ядра обнаружен вход от зрительных областей новой коры (рис. 77) и их аналога в мозге птиц. Наконец, общее заключается в существовании входов от тектума. Сходство их эфферентов заключается в установлении связей с ядрами глазодвигательного комплекса, хотя они выражены по-разному у разных представителей. Так, сложно организованные связи описаны между добавочной зрительной системой и глазодвигательным комплексом у'птиц. Строго упорядоченные проекции имеются также у амфибий, телеостей и рептилий (рис. 78). У млекопитающих не описаны прямые связи добавочной зрительной системы и ядер глазодвигательного комплекса, что же касается претектума, то его структуры проецируются на ядро отводящего нерва (прямо или через медиальную ретикулярную формацию) у низших млекопитающих, у высших же таких связей не обнаружено.
Отсутствуют они и у среднемозгового чечевицеобразного ядра птиц.
Другое направление эфферентов связывает ядра обеих систем с вестибуло-латеральной долей мозжечка. Причем, у высших позвоночных к прямым входам в мозжечок добавляются и опосредованные нижней оливой, а у млекопитающих описаны исключительно опосредованные (нижней оливой и ядрами моста) связи. У птиц, черепах и некоторых млекопитающих имеются связи добавочной зрительной системы с вестибулярным комплексом, а у птиц и млекопитающих - и с ядрами Кахаля и Даркшевича. Претектальная область таких входов не образует.
Анализ распределения связей и изучение характеристик нейронной активности претектальной области и добавочной зрительной системы позволили установить, что эти образования играют решающую роль в организации зрительно-моторных реакций; в частности, опосредуют вертикальный и горизонтальный нистагмы, а кроме того, принимают участие в стабилизационных процессах на сетчатке. Наконец, связи оливарного претектального ядра с ядром Эдингера - Вестфаля обусловливают зрачковый рефлекс. Компоненты обеих систем, имеют тесные связи между собой, а также с тектумом. Однако возможно, что сказанное не в полной мере отражает функциональную роль претектальной области, элементы которой, по крайней мере у птиц, некоторых рептилий и у высших млекопитающих, проецируются на филогенетически молодые таламические структуры и, вероятно, опосредуют проведение зрительной информации в теленцефалон.
* Отметим, что у примитивных млекопитающих таламус часто представлен относительно гомогенной клеточной массой, лишенной внутренних топографических ориентиров, что затрудняет его деление (рис. 73).
* Отметим, что в составе данной группы у приматов описывают и дополнительные ядра: латеральное промежуточное и, в составе LP, - угловое.
§ 2. ОБЩАЯ ХАРАКТЕРИСТИКА ГИПОТАЛАМУСА
Гипоталамус, Hypothalamus, занимает самое вентральное положение среди диенцефальных образований. Он представлен нервной тканью, окружающей нижнюю часть III желудочка и сосредоточенной в основании мозга. Относительно малые размеры этого отдела, отсутствие крупных миелинизированных трактов, диффузное распределение нейронов, составляющих непрерывную клеточную массу с различной степенью концентрации элементов, - все эти черты характерны для гипоталамуса позвоночных независимо от их таксономической принадлежности и обусловливают объективные сложности классификации гипоталамических образований даже в пределах одной группы.
Наиболее часто структуры гипоталамуса разделяют (даже у млекопитающих) на зоны в зависимости от их удаленности от вентрикулярной стенки: перивентрикулярную, медиальную и латеральную. Степень развития разных отделов гипоталамуса варьирует: у многих низших позвоночных основные клеточные элементы сосредоточены в перивентрикулярной области, у высших представителей отдельных таксономических групп пластиножаберные,
костистые рыбы, рептилии) дифференциация структур гипоталамуса идет прежде всего за счет медиальной зоны, а у птиц и млекопитающих - за счет латеральной.
В составе гипоталамуса часто выделяют в качестве самостоятельного отдела преоптшескую область (АР), занимающую наиболее ростральное положение в этом отделе мозга (рис. 67). Ее часто рассматривают как неэвагинировавшую часть конечно-то мозга. Особенно хорошо данная область выражена у низших позвоночных. У низших млекопитающих в ней выделяют те же зоны, что и в остальной части гипоталамуса: латеральную, медиальную и перивентрикулярную. У приматов данная область дифференцирована крайне слабо.
У низших позвоночных гипоталамус образован тонкостенной структурой, лежащей в основании мозга, конфигурация которой зависит от формы III желудочка - многочисленные углубления вентрикулярной полости приводят к изменению очертаний этого отдела и вместе с перекрестом зрительного нерва часто служат ориентирами для разделения гипоталамуса на области или отделы (рис. 65, 67).
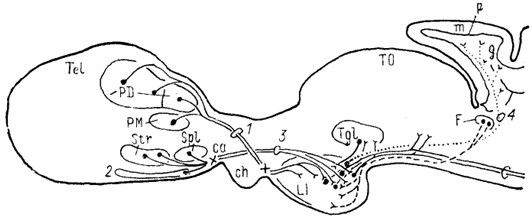
Рис. 79. Основные связи нижней доли гипоталамуса ската Raja eglanteria (Smeets, Boord, 1985).
1 - паллиальный тракт, 2 - базальная поверхностная область, 3 - МПП, 4 - лобо-церебеллярный тракт. X - перекрест волокон.
У многих круглоротых основные клеточные группы сосредоточены вблизи желудочка. У миноговых, хрящевых и лучеперых рыб в составе гипоталамуса появляются образования, удаленные от вентрикулярной стенки. Специфическим отделом мозга этих животных является крупная нижняя доля гипоталамуса, lobus inferior hypothalami.
Нижняя доля - структура гетерогенная, включающая несколько клеточных групп (рис. 66). Данные относительно ее связей фрагментарны. Показано, что у рыб с высокодифференцированной вкусовой системой этот отдел мозга получает входы от вторичного вкусового ядра или от его гомолога-ядра F (у пластиножаберных рыб). Другие афференты поступают от тегментальной области и тектума среднего мозга, мозжечка, а у пластиножаберных рыб - от контрлатерального полушария конечного мозга (рис. 79). Эфференты нижней доли у пластиножаберных
и костистых рыб направляются к моторным структурам ствола мозга. В последнее время подтверждено существование у пластиножаберных рыб связей нижней доли с мозжечком - это лобо-церебеллярный тракт, описанный ранее. Он начинается в каудальной части нижней доли и адресуется передней части тела мозжечка. Физиологические данные свидетельствуют об участии нижней доли в регуляции пищевого поведения у скатов.
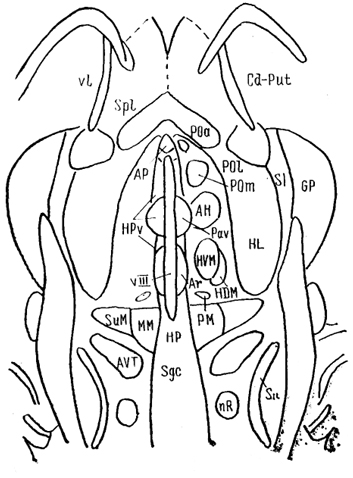
Рис. 80. Ядра гипоталамуса мозга крысы на горизонтальном срезе (Swanson, 1987).
АН - переднее гипоталамическое, Аг - аркуатное, РМ - премамиллярное, ММ - мамиллярное, SuM - супрамамиллярное, РОа - переднее, Pol - латеральное и Рот - медиальное преоптические ядра, SI - безымянная субстанция.
Остальные отделы гипоталамуса рыб во многих отношениях сходны с таковыми наземных позвоночных различия заключаются лишь в степени развития каждой из зон.
У всех позвоночных перивентрикулярная зона гипоталамуса (HPv) организована довольно просто в состоит из примитивных клеток изодендритного типа. Нейроны образуют узкую клеточную полоску примыкающую к стенке желудочка и пронизанную многочиеленными волокнами перивентрикулярной системы. Наиболее заметной клеточной агрегацией этой зоны является паравентрикулярное ядро (nucl. paraventricularis), которое сохраняет свое положение и в мозге других позвоночных (рис. 69, 80).
Общая черта всех позвоночных - это наличие в данной зоне нейросекреторных клеток, а также нервных элементов, получающих прямой вход от волокон зрительного нерва. Сама зона тесно связана со структурами циркумвентрикулярной системы в области III желудочка (в частности, у низших позвоночных здесь широко представлены клетки-таннициты, расположенные в срединном возвышении и способные к активному транспорту и абсорбции нейрогормонов). Число нейросекреторных клеток особенно велико у низших позвоночных в преоптической перивентрикулярной области, а у наземных - в супраоптическом и: паравентрикулярном ядрах. Их аксоны образуют гипоталамо-гипофизарный тракт и выделяют нейрогормоны и рилизинг-факторы
в отделы гипофиза. Тем самым обеспечивается; прямая связь мозга с эндокринной системой.
Другой чертой, характерной для перивентрикулярной зоны гипоталамуса, является наличие клеток-мишеней зрительных афферентов. Зрительные проекции адресованы, и передней части) медиальной зоны. Они описаны у всех исследованных видов, позвоночных, но районы их окончания обозначаются по-разному. У хрящевых рыб структура, получающая такие входы, обозначалась разными авторами как гипоталамическое зрительное ядро, хиазматическое ядро, супрахиазматическое ядро.* Под последним, наиболее распространенным, названием оно описано у многих представителей позвоночных. Полагают, что у всех позвоночных эта структура гипоталамуса связана с регуляцией циркадной ритмики различных процессов в. организме, в том числе циклов сон - бодрствование - у высших. У млекопитающих помимо входа от сетчатки сюда адресованы проекции ЛКТ, а эфференты связывают это ядро с эпифизом, оказывая влияние на продуцирование последним гормонов.
Таким образом, перивентрикулярная зона гипоталамуса имеет сходное строение у всех позвоночных, в основном содержит нейроны, обеспечивающие связи гипоталамуса с эндрокринными органами и обусловливает регуляцию циклических процессов, зависящих от степени освещенности.
У наземных позвоночных наиболее обширна медиальная зона гипоталамуса (НМ). Она состоит из нескольких ядер, частей разделяемых в соответствии с расположением в рострокаудальном направлении на ядра передней, средней и задней групп. Ядра передней группы (SO, Sch) содержат нейросекреторные клетки и нейроны, получающие зрительные афференты из сетчатки, и оказываются в структурном и функциональном отношениях сходными с элементами перивентрикулярной зоны. В средней группе основными являются вентро- и дорсомедиальное ядра гипоталамуса (HVM, HDM). Задняя группа (HP) слабо выражена у низших позвоночных и только намечается у хвостатых амфибий. Вообще у анамний и рептилий супраоптическое и супрахиазматическое ядра имеют диффузное строение, а зона НМ слабо намечена. При этом некоторые ядра (например, супраоптическое) присутствуют почти у всех позвоночных, другие (например, некоторые ядра задней группы) дифференцируются только у млекопитающих.
Основной вход в ядра медиальной зоны (за исключением передней группы) составляют отделы лимбической системы конечного мозга (амигдала, септальная область), а также сенсорные ядра ствола мозга, обеспечивающие проведение информации от вкусовых, висцеральных, обонятельных, зрительных, а
у млекопитающих и от соматических и слуховых рецепторов. Эфференты адресованы соседним, перивентрикулярной и латеральной, зонам гипоталамуса, а также - в составе медиального пучка переднего мозга - лимбическим структурам, ретикулярной формации, району центрального серого вещества среднего мозга и, наконец, премоторным и вегетативным ядрам ствола.
В функциональном отношении охарактеризовать структуры медиальной зоны довольно сложно. Очевидно, что их совокупная деятельность обеспечивает участие этой области промежуточного мозга в организации сложных форм пищевого, питьевого, полового и т. п. поведения, что обусловлено как наличием разнообразных сенсорных входов, так и множественными выходами на исполнительные системы организма (через перивентрикулярную зону - на эндокринную систему, через премоторные и вегетативные ядра - на сомато- и висцеромоторные образования) и к лимбическим структурам конечного и среднего мозга.
Среди всех вышеназванных структур медиальной зоны выделяются мамиллярные тела, corpora mammillares, которые хорошо выражены у высших позвоночных. У млекопитающих они представлены крупными выпуклыми образованиями, лежащими на вентральной поверхности мозга и состоящими из нескольких ядер. Система их связей оказывается иной, чем у других ядер медиальной зоны. Основной вход к мамиллярным телам образован гиппокампальными нейронами, аксоны которых у млекопитающих образуют крупный проводящий тракт - свод, fornix. Сенсорные афференты, в отличие от других ядер, приходят сюда из сенсорных полей (зрительной и слуховой областей) новой коры. Характер распределения эфферентов также различается: основные проекции адресованы ядрам передней группы таламуса, а через них - лимбической области коры. Система внутриги-поталамических связей у мамиллярных тел не развита. Полагают, что мамиллярные тела и связанные с ними образования имеют отношение к организации процессов обучения и памяти.
Необходимо отметить, что у млекопитающих в составе мамиллярного комплекса часто описывают группу клеток, которая в ходе эволюции этого класса позвоночных вычленяется в самостоятельный крупный комплекс - ядра серого бугра, или туберальные ядра. Слабо выраженный у низших млекопитающих, он значительно эволюционирует среди приматов, а у антропоидов и человека имеет в своем составе ядра, отсутствующие у других приматов. Учитывая тесные связи данных ядер с корой, полагают, что развитие этого отдела шло параллельно с развитием некоторых отделов новой коры.
Нейронные группы, лежащие среди волокон медиального пучка переднего мозга, составляют латеральную зону гипоталамуса (HL). У птиц и млекопитающих она достигает значительных размеров и дифференцируется на ряд полей и ядер. У антропоидов в ее пределах дополнительно выделяется латеральная
преоптическая зона, латеральное и супрамамиллярное ядра, отсутствующие у других высших позвоночных. В силу особенностей строения и связей эту зону часто рассматривают как .диенцефальный компонент ретикулярной формации. Об этом же свидетельствуют результаты физиологических наблюдений, в частности стимуляция этой области приводит к реакции активации, характерной для восходящей ретикулярной формации. Относительно афферентов этой области сведения немногочисленны. Эфференты распределяются почти по всей поверхности новой коры, в лимбических структурах конечного мозга, а также в моторных и вегетативных ядрах ствола мозга.
Основные связи гипоталамуса сходны у всех позвоночных. Большинство афферентов и эфферентов этой области проходит в составе многокомпонентного медиального пучка переднего мозга (МППМ, fasciculus prosencephali medialis). В мозге млекопитающих он связывает более чем 50 образований, начиная от ростральных отделов конечного мозга и кончая спинным. Значительное число эфферентов входит в состав перивентрикулярной системы волокон, связывающих гипоталамус с таламусом, средним мозгом, перивентрикулярно расположенными отделами ствола. Собственной системой связей гипоталамуса является гипоталамо-гипофизарный тракт, реализующий воздействие на эндокринную систему. Связи со структурами старой коры образованы у наземных позвоночных специфической системой волокон - сводом.
Таким образом, во многих отношениях гипоталамус оказывается консервативным отделом мозга, сохраняющим сходные черты организации у всех позвоночных. Наиболее ярко это проявляется в строении и системе связей перивентрикулярной зоны. Вместе с тем в гипоталамусе возникают черты прогрессивной дифференцировки, которая выражается у высших рыб в развитии нижней доли, а у птиц и млекопитающих - латеральной зоны. В то же время у других представителей позвоночных (низшие лучеперые рыбы, двоякодышащие, амфибии) сохраняется исходное, перивентрикулярное положение основных элементов гипоталамуса.
Несмотря на многочисленные исследования различия в подходах не позволяют провести гомологизацию многих отделов гипоталамуса разных позвоночных. Однако в тех случаях, когда это возможно, четко прослеживаются черты прогрессивных преобразований в ряде отделов гипоталамуса у млекопитающих по сравнению с птицами, а среди млекопитающих - у приматов по сравнению с другими группами. Это выражается прежде всего в появлении у приматов в пределах задней группы ядер медиальной зоны дополнительных объединений и в дифференциации латеральной зоны (при сравнении с птицами). На нейронном уровне прогрессивные тенденции в развитии гипоталамуса сказываются в появлении у некоторых млекопитающих
тенденции к сосредоточению нейронов в пространственные комплексы-кластеры.
В целом же в составе гипоталамуса выделяют аналогичные области, элементы которых, имея сходные связи, вероятно, выполняют и сходные функции у разных позвоночных. Это, во-первых, обилие выходов на эндокринную систему, реализуемых через гипофиз и в основном осуществляемых элементами перивентрикулярной области. Во-вторых, конвергенция разномодальной импульсации (у низших, в основном, висцеросоматической, а у высших и экстероцептивной) и множество доступов к исполнительным отделам мозга (моторным и вегетативным ядрам), обеспечивающие участие гипоталамуса в организации поведенческих реакций, в том числе сложных комплексных форм поведения (у скатов, птиц и млекопитающих). В-третьих, наличие у всех позвоночных ретинальных входов в гипоталамус, что, вероятно, представляет собой реализацию древнего механизма регуляции жизнедеятельности организма в зависимости от степени освещенности и времени суток и осуществляется за счет влияния гипоталамуса на гипофиз, а у высших позвоночных и на эпифиз.
Наконец, согласно физиологическим данным, в гипоталамусе млекопитающих обнаружены так называемые "центральные" рецепторы, активация которых возможна не синаптическим путем, а сенсорными стимулами (центральные глюко- и терморецепторы). В ходе эволюции происходит дифференциация структур гипоталамуса, приводящая к формированию участков, связанных с высшими отделами мозга и вовлеченных таким образом в регуляцию и обеспечение сложных форм поведенческих реакций. Отметим, что у млекопитающих гипоталамус рассматривается в качестве "выходного" звена лимбической системы именно с учетом характера его связей.
* У хрящевых рыб Platyrhinoidis зрительный вход- обнаружен в каудальном гипоталамусе на уровне заднего бугорка.
§ 3. ЭПИТАЛАМУС
Эпиталамус, Epithalamus, состоит из эпифиза (шишковидная железа, верхний мозговой придаток, Epiphysis) и поводковых (хабенулярных) ядер, nucll. habenulae.
Эпифиз в эволюции позвоночных развивался на основе так называемого теменного глаза низших позвоночных, который сформировался как производное одного или двух выростов дорсальной стенки промежуточного мозга - собственно теменного (парапинеального) органа, расположенного рострально, и пинеального органа, занимающего каудальное положение. Оба эти образования могут быть одинаково развитыми у одного и того же животного и содержать светочувствительные элементы (панцирные и костистые рыбы). У других позвоночных преобладающей структурой является пинеальный орган (круглоротые) или теменной (гаттерии, некоторые ящерицы). У высших позвоночных
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|