- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 15 страница
связи рассматриваются как основные, по крайней мере в мозге низших позвоночных. Так, у круглоротых нейроны области шва и лежащие в ее пределах мюллеровские клетки охватывают своим влиянием весь спинной мозг. В мозге рыб сохраняются гигантские клетки, а основные нисходящие связи также сформированы ретикулярными ядрами. В мозге высших позвоночных гигантские клетки отсутствуют, однако у них сохраняются тракты, происходящие из ретикулярных ядер. Сравнение нейронного состава источников нисходящих трактов, путей их прохождения в спинном мозге, особенностей медиаторного обеспечения позволили гомологизировать те отделы, которые дают начало ретикуло-спинальным волокнам.
Некоторые нисходящие тракты происходят из рано выделившихся в ходе эволюции объединений ретикулоподобных нейронов (ядра МПП, ядра Кахаля, а у высших и ядра Даркшевича) и, следовательно, также относятся к ретикуло-моторным. Отметим, что в результате эволюционных преобразований источники ретикуло-моторных связей получают все больший объем афферентов от вышележащих отделов мозга и включаются в новые моторные системы более высокого порядка. Так, у млекопитающих многие волокна кортико-бульбарного тракта заканчиваются на каудальных отделах ретикулярной формации.
Усложнение общей организации мозга приводит к появлению связанных с обеспечением локомоции областей, которые совпадают с определенными отделами ретикулярной формации, - среднемозговая локомоторная область, совпадающая с областью клиноподобного ретикулярного ядра у млекопитающих, ее эквиваленты в тегментуме костистых и пластиножаберных, области, осуществляющие иннервацию электромотонейронов у электрических рыб.
Ретикулярная формация дает начало многим ядерным группам мозга высших позвоночных - верхнеоливарному комплексу, трапециевидному телу, красному ядру, которые постепенно обособляются из ретикулярного окружения. Таким образом, можно полагать, что большинство отделов надсегментарного уровня являются дериватами ретикулярной формации. Кроме того отметим, что у большинства позвоночных в пределах ретикулярной формации локализуются нейроны, осуществляющие иннервацию рецепторных отделов разных сенсорных систем.
Области ретикулярной формации составлены клетками, которые являются местом конвергенции разномодальной импульсации и имеют множественные выходы на моторные элементы мозга. Следовательно, они соответствуют представлениям о сущности интегративных отделов мозга, и в этом смысле ретикулярная формация может рассматриваться как древнейшая интегративная система мозга позвоночных, в строении элементов которой есть ряд специфических черт, оказавшихся универсальными и сохранившихся и в других отделах мозга.
Несмотря на множество исследований, посвященных ретикулярной формации разных позвоночных, многие вопросы далеки от своего разрешения. Это относится и к созданию единой номенклатуры, и к критериям разделения ретикулярных образований на зоны, и к возможности и правомочности выделения ретикулярных участков в промежуточном и конечном, а также спинном мозге. Вероятный прогресс в решении этих вопросов связывают с использованием нейрохимического подхода и с разработкой функциональных критериев.
Ретикулярная формация представляет собой средоточие интернейронов. В ходе эволюции достаточно рано в ее составе вычленяются локальные клеточные группы, дающие начало ядрам ретикулярной формации. Направления эволюционных преобразований этого отдела ЦНС, вероятно, различны. Наиболее четко уже у низших позвоночных выражено формирование участков, обеспечивающих поступление разномодальной информации к моторным отделам мозга; в их пределах часто обнаруживаются особые гигантские нейроны (нейроны Мюллера у бесчелюстных и маутнеровские клетки у рыб). Вместе с тем некоторые зоны ретикулярной формации содержат клетки, обеспечивающие взаимосвязи многих отделов мозга в результате распространения их аксонов на относительно большие расстояния. Так, уже у бесчелюстных можно выделить ядра области шва, содержащие серотонинергические нейроны с проекциями, сходными с имеющимися у высших. Наконец, в пределах ретикулярной формации у высших позвоночных формируется так называемая ретикулярная активирующая система, и хотя вопрос о ее гомологах в мозге других позвоночных окончательно не решен, возможно, что это результат развития восходящих проекций медиаторноспецифичных отделов ствола. Бесспорным на сегодня является рассмотрение ретикулярной формации как первой в эволюции интегративной системы мозга, поскольку именно здесь конвергируют проекции разных сенсорных систем, а связи ретикулярных ядер с моторными обусловливают активацию исполнительных отделов мозга, обеспечивая тем самым протекание адаптивных реакций организма.
ГЛАВА 7
ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг, Diencephalon, развивается рострально от ствола мозга и претерпевает в ходе эволюции значительные преобразования, во многих отношениях связанные с совершенствованием теленцефальных отделов.
Структурная организация промежуточного мозга исследовалась с помощью разных методик и на множестве представителей позвоночных. Тем не менее до сих пор остаются нерешенными вопросы гомологии не только компонентов, но и целых отделов промежуточного мозга. Традиционно диенцефальные образования делятся на эпиталамус, гипоталамус и собственно таламус (другие отделы описаны для конкретных групп животных). В качестве их границ многие авторы рассматривают борозды на вентрикулярной поверхности, что, по их мнению, позволяет выделить гомологичные отделы на основании сходства их закладки в эмбриогенезе. Однако объективность и полнота подобного подхода неоднократно оспаривались и в последние десятилетия предприняты попытки введения дополнительных, а в некоторых случаях иных, критериев для разделения промежуточного мозга.
Итак, вопрос оказывается далеким от разрешения, что в первую очередь относится к мозгу анамний. У них в силу перивентрикулярного расположения основной массы клеточных элементов часто проводят разделение диенцефалона по зонам, расположенным параллельно оси мозга, и таким образом выделяют перивентрикулярные, наружные перивентрикулярные и мигрировавшие в толщу мозга элементы. Часто этот подход используют в дополнение к традиционному, предполагая зональную характеристику каждого из отделов. Кроме того, в мозге низших позвоночных описывают отделы, отсутствующие у высших. Дополнительные разночтения возникают при решении вопроса о принадлежности структур, занимающих пограничное положение.
§ 1. СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ТАЛАМУСА
Таламус, Thalamus, или зрительный бугор, - отдел промежуточного мозга, который претерпевает наибольшие структурные преобразования в ряду позвоночных, что во многом обусловлено его взаимосвязями с прогрессивно развивающимися отделами конечного мозга. Многонаправленность преобразований, недостаточность фактических сведений экспериментальной морфологии, а также неразработанность общих критериев разделения? таламических структур обусловили огромное разнообразие классификаций, а также противоречивость мнений относительно гомологичности отдельных структур таламуса.
Строение таламуса низших позвоночных
Традиционно в пределах таламуса всех позвоночных выделяют дорсальный и вентральный отделы (ThD, ThV). У круглоротых в состав дорсального таламуса входят расположенные перивентрикулярно слоистое серое вещество, в последнее время обозначаемое как дорсальное таламическое ядро (Thd), а также наружное коленчатое тело (CGL), в состав которого включены описанные ранее как самостоятельные ядро Беллончи; (nВ), переднее и заднее коленчатые ядра. Общими для всех названных образований являются ретинальные входы. Кроме того, пВ получает проекции обонятельных отделов конечного мозга, гипоталамуса и тектума. К переднему коленчатому ядру подходят спинно- и бульбо-таламические тракты, к заднему - проекций октавного комплекса. Эфференты связывают дорсальный таламус с продолговатым и средним мозгом (тегментальная область) и с гипоталамусом. Показан также восходящий путь- tr. thalamofrontalis, - возможно, адресованный гиппокампу.
Вентральный таламус у миног отделен от дорсального разреженной зоной (см. рис. 29) и состоит из вентрального таламического ядра, nucl. ventralis thalami (Thv), и ядра заднего бугорка, nucl. tuberculi posterioris (TP). Эфференты последнего, присоединяясь к медиальному продольному пучку, направляются в спинной мозг. Небольшое количество волокон происходит из вентрального ядра.
У хрящевых рыб как в дорсальном, так и в вентральном таламусе выделяют медиальную и латеральную части (рис. 65, 66). Медиальная представлена перивентрикулярно расположенными плотно лежащими мелкими нейронами (рис. 65, а), латеральная-мигрировавшими от вентрикулярной стенки, крупными элементами (рис. 65, б).
Наиболее крупным отделом дорсального таламуса пластиножаберных является так называемое латеральное коленчатое тело (ЛКТ). Однако несмотря на идентичность названий доказательства его гомологичности одноименному ядру млекопитающих отсутствуют. В связи с этим ряд авторов предпочитают использовать
иное название, другие рассматривают данную структуру как комплекс, состоящий из нескольких отделов. Так, у Ginglymostoma лишь дорсальную часть "ЛКТ" описывают как зрительную, сохраняя за ней это название (другие называют эту часть дорсолатеральным оптическим комплексом); вентральную же часть называют вентролатеральным оптическим ядром. При этом вход сетчатки адресован только последнему, а к дорсальной части поступают лишь тектальные афференты. Эфференты обеих нейронных групп направляются в паллиум. Согласно существующим представлениям вентролатеральное оптическое ядро можно рассматривать в качестве гомолога вентральному ЛКТ, а дорсальную группу - как гомологичную комплексу "дорсальное ЛКТ - LP" млекопитающих.

Рис. 65. Отделы промежуточного мозга в проекции на вентрикулярную поверхность III желудочка у акулы Scyliorhinus canicula(Smeets e. a., 1983).
а, б - группы ядер: перивентрикулярно лежащие (а), удаленные от вентрикулярной стенки (б). 1 - поперечный парус. Штриховая линия - ядра нижней доли гипоталамуса.
Центральное таламическое ядро большинства видов лишено сетчаточных входов, но получает проекции тектума и спинного мозга. Ядро, описанное как латеральное заднее, имеет афференты
от мезенцефалического центра представительства электросенсорной системы. Окончания других центров октаво-латеральной системы в таламусе хрящевых рыб не исследованы. Вентральный таламус представлен клеточными массами, формирующими нисходящие проекции к спинному мозгу. Относительно его .афферентов сведения отсутствуют.
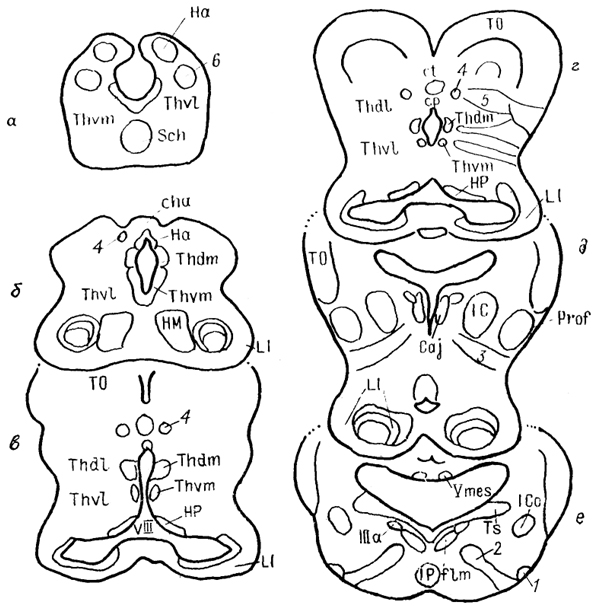
Рис. 66. Строение промежуточного мозга и рострального отдела среднего мозга акулы Scyliorhinus canicula (Reperant e. a., 1986).
1-3 - структуры добавочной оптической системы: базальный зрительный тракт (1), зрительное ядро вентрального тегментума среднего мозга (2), зрительная область, дорсального тегментума среднего мозга (3); 4, 5 - претектальные ядра: оптическое ядро задней комиссуры (4), оптическое центральное ядро (5), 6 - дорсолатеральное ядро, а-e - срезы в ростро-каудальном направлении.
В мозге актиноптеригий в составе таламуса помимо дорсального и вентрального отделов в качестве самостоятельной области выделяют задний бугорок, pars tuberculi posterioris (TP) (рис. 67, 1). Он весьма значительно развит у этих животных и вопрос о его гомологичности отделам таламуса других позвоночных не решен. В состав заднего бугорка входит значительное число нейронных групп. В пределах перивентрикулярной области располагаются ядро заднего бугорка, перивентрикулярное
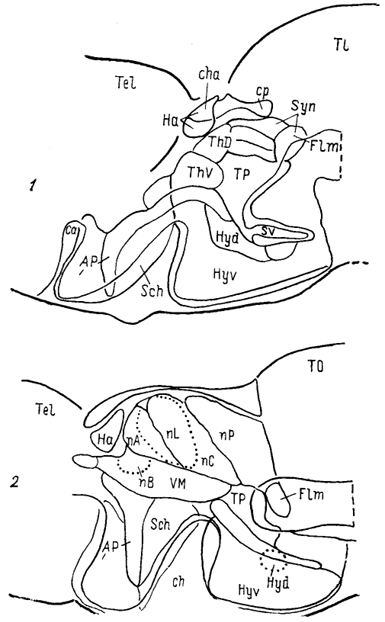
Рис. 67. Схема организации промежуточного мозга представителей многоперовых рыб - Polypterus palmas (1) и бесхвостых амфибий Rana catesbeiana (2) в проекции на вентрикулярную поверхность III желудочка (Braford, Northcutt, 1983).
Тонкие линии - контуры ядер, жирные - контуры мозга.
ядро заднего бугорка, паравентрикулярный орган и его ядро. Мигрировавшие клеточные массы образуют разное количество групп, среди которых наиболее постоянными являются гломерулярное и прегломерулярное ядра, nucll. glomerulosus et preglomerulosus (Gl, pGl). У большинства представителей ядро Gl - крупная сферическая клеточная масса с четким гломерулярным нейропилем. Ранее оно описано как круглое ядро, nucl. гоlundus, однако используемое в настоящее время название позволяет не только избежать отождествления его с круглым •ядром других позвоночных (что вовсе не подтверждено), но и избавиться от разночтений, поскольку "круглым ядром" разные авторы называли структуры и таламуса и преоптической области лучеперых. Так, ядро G1 отсутствует у карповых, а название "круглое ядро" дано у них структуре, которая значительно отличается от гломерулярного ядра наличием выраженных входов от вторичного вкусового ядра. Прегломерулярное ядро чаще всего рассматривается как комплекс ядер.
Дорсальный таламус лучеперых рыб включает перивентрикулярно лежащее переднее ядро, nucl. anterior thalami (nA), a также дорсальное и центральное задние ядра, nucll. dorsalis posterior et centralis posterior (DP, CP). Вентральный таламус состоит из таламического возвышения, eminentia thalami (ETh) и лежащих каудально от него промежуточного, вентролатерального и вентромедиального ядер, соответственно VI, VL, VM (рис. 68). Соотношение основных отделов таламуса у разных
групп лучеперых оказывается различным, наибольшие вариации касаются области заднего бугорка.
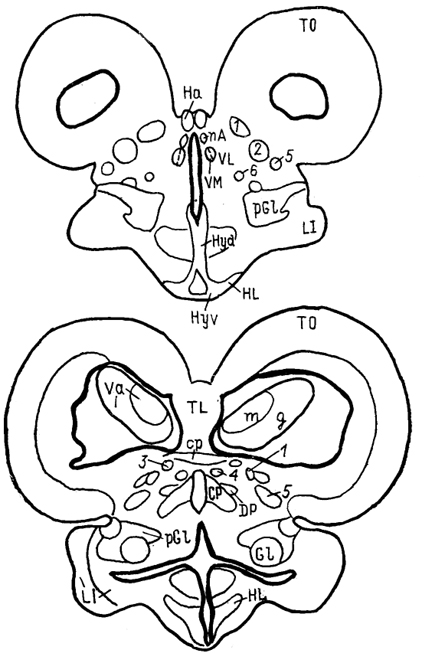
Рис. 68. Ядра промежуточного мозга Carassius auratus (no: Wullimann, Northcutt, 1988).
1-4 - претектальные ядра: центральное (1), поверхностное, крупноклеточная часть (2), 3, 4 - перивентрикулярное: дорсальная (3) и вентральная (4) части; 5, 6 - ядра добавочного зрительного тракта: дорсальное (5) и вентральное (6).
Характер связей таламуса лучеперых рыб исследован недостаточно полно. Наиболее многочисленны сведения относительно входов зрительного тракта, которые обнаружены у представителей всех групп. Нейроны, получающие контрлатеральный ретинальный вход, локализованы в пределах переднего таламического ядра, латерально от промежуточного и вблизи вентролатерального. Ипсилатеральные входы организованы различно. Так, у хондростей и голостей они адресованы структурам, имеющим
и контрлатеральные входы. На основании сходства в их распределении с таковым у высших позвоночных полагают, что это же было свойственно древним актиноптеригиям, у ранних костистых рыб такие связи были утрачены, а у ныне живущих видов они оказались приобретенными вторично, независимо от других представителей. Действительно, у некоторых телеостей ипсилатеральные входы сетчатки в таламус отсутствуют, у других они возникают в результате двойного перекреста и адресуются ростральным отделам.
Сведения о других афферентах весьма фрагментарны. Так, у некоторых исследованных в этом отношении видов, представляющих основные группы лучеперых рыб, обнаружены входы обонятельных луковиц в область заднего бугорка. Тектальные проекции есть у всех ядер, получающих ретинальные афференты, а также у ядер DP, CP, VM; при этом они показаны и у слепых пещерных рыб (в частности, у Astyanax hubbsi - в область "ЛКТ"). Проекции торуса у голостей направляются в ядро СР и прегломерулярный комплекс, однако детали представленности входов от вестибулярного комплекса и органов боковой линии остаются неясными. У костистых рыб проекции боковой линии подходят к области вентрального таламуса, получающей афференты и от латеральной части теленцефалона.
Местом окончания церебелло-диенцефальных путей у голостей и телеостей являются ядра VM, nA, TP, при этом основной объем входов адресован VM. Область заднего бугорка получает также восходящие проекции вторичного вкусового ядра и - у телеостей - конечного мозга.
Эфференты таламуса лучеперых рыб практически не исследованы. Имеются сведения о восходящих в теленцефалон проекциях от pGl. Дорсальные ядра образуют связи с тектумом, G1- с моторными ядрами V и VII черепномозговых нервов, область заднего бугорка - с долей лицевого нерва.
Помимо названных отделов в таламус лучеперых рыб включают так называемый латеральный валик, torus lateralis (Tla). У многоперовых, хондростей и голостей он представляет собой крупное вытянутое скопление нейронов, расположенных на латеральной поверхности в каудальной части промежуточного мозга. У костистых рыб эта структура значительно меньше и смещена рострально. Связи, происхождение и возможные гомологи этой структуры неясны; некоторые авторы считают ее связанной с продольным торусом.
Следует отметить, что в числе образований промежуточного мозга у костистых рыб ряд авторов выделяет в качестве самостоятельного еще один отдел - synencephlon (Syn), который занимает пограничную область между промежуточным и средним мозгом (рис. 67, 1). В состав этого отдела включают дорсально расположенную область задней комиссуры, regio com-missurae posterioris, и лежащее вентрально ядро медиального
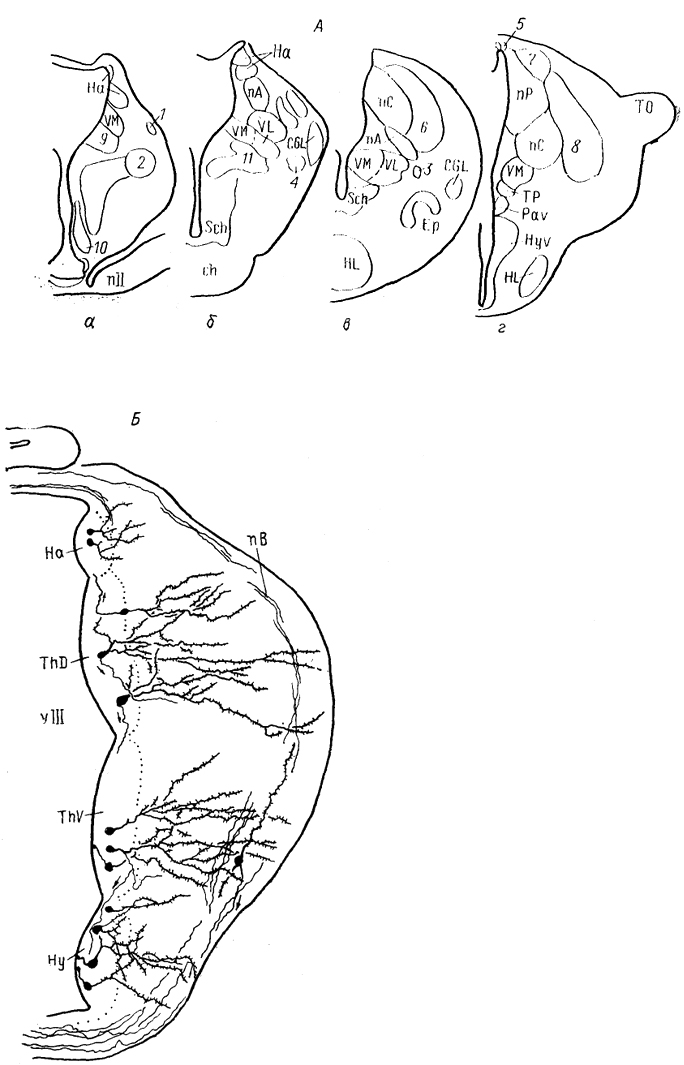
Рис. 69. Схема организации промежуточного мозга представителей бесхвостых амфибий JRana catesbeiana (А) и хвостатых амфибий - Нупоbius (Б) (Neary, Northcutt, 1983; Обухов, 1990).
1 - ядро ложа медуллярной полоски; 2 - латеральная амигдала; 3 - нейропиль Беллончи; 4 - поверхностное желудочковое ядро вентрального таламуса; 5 - субкомиссуральный орган; 6 - 8 - части латерального ядра: передняя (6), дорсальная (7) и вентральная (8); 9-11 - части преоптического ядра; дорсальная (9), вентральная (10) и крупноклеточная (11). а-г - срезы в рострокаудальном направлении.
продольного пучка (у большинства позвоночных область задней комиссуры относят к претектальной области).
Таким образом, в таламусе водных позвоночных можно выделить, насколько нам позволяют имеющиеся данные, группу ядер, являющихся диенцефальным представительством сенсорных систем. Наибольшее число сведений касается зрительной: системы, однако изучение других сенсорных входов также свидетельствует о наличии на этом уровне центров представительства иных модальностей. Вместе с тем, отсутствие информации о распределении теленцефальных эфферентов и недостаток экспериментально-морфологических исследований не позволяют проводить корректные аналогии с организацией таламуса наземных позвоночных.
В мозге амфибий в последнее время также разделяют мигрировавшие и перивентрикулярно расположенные структуры (рис. 69), кроме того, некоторые авторы описывают в пределах таламуса и область заднего бугорка (рис. 67, 2).
Дорсальный таламус бесхвостых амфибий состоит из латерального ядра (L) и перивентрикулярно расположенных переднего (nА), центрального (nС) и заднего (nР) ядер (рис. 69, A); двум последним свойственно четкое ламинарное строение. Исходя из характера связей дорсальный таламус делят на три зоны: переднюю, среднюю и заднюю.
Передняя зона включает лишь nА. Она получает проекции от медиального паллиума, вентрального гипоталамуса, энтопедункулярного ядра, области задвижки продолговатого мозга, а также (за счет ветвления дендритов собственных нейронов в областях окончания ретинальных афферентов и среднемозговом акустическом центре) от структур зрительной и слуховой систем. Входящее в состав передней зоны nА является единственным (по имеющимся на сегодня сведениям) источником таламических входов в медиальный паллиум и вентральную часть дорсального паллиума. Вполне вероятно, что нейроны, дающие начало этим эфферентам, локализованы в одной И той же части ядра, но вопрос о том, являются ли эти клетки источниками обоих входов или же те происходят из соседних нейронных групп, остается невыясненным. Отметим, что таламо-теленцефальные проекции, о которых идет речь, билатеральны с преобладанием ипсилатерального звена. Учитывая наличие разно-модальных сенсорных входов в пА, его можно рассматривать как важный (а может быть и единственный) центр проведения сенсорной информации в паллиальные отделы мозга бесхвостых амфибий. Другие эфференты передней зоны адресованы вентральному гипоталамусу, тектуму, задней зоне таламуса.
Средняя зона, более крупная, состоит из nC, а также переднего и задневентрального отделов латерального ядра (La, Lpv). Структуры средней зоны получают входы от тектума (La, Lpvy и торуса (nС), кроме того, имеются соматические сенсорные
проекции от областей среднего мозга, обладающих соматотопической организацией, и от области задвижки. Основные эфференты этой зоны адресованы стриатуму, и таким образом она рассматривается в качестве важного звена проведения гетеромодальной информации в вентролатеральные области конечного мозга. Небольшое число эфферентов nС направляется в тектум, a Lpv - в торус.
Задняя таламическая зона включает в себя ядро nР и заднедорсальную часть латерального ядра, Lpd. Она получает проекции от других диенцефальных областей (передней зоны дорсального таламуса, вентрального гипоталамуса, пинеального органа), стриатума, тектума, кроме того, дендриты ее нейронов, проникая в соседние области, получают ретинальные входы. Эфференты зоны не проецируются в конечный мозг, но адресуются множеству других отделов: моторным ядрам ствола и спинного мозга, тектуму, а также (лишь от ядра nC) торусу, вентральному таламическому ядру VL и nА.
Гомологизация ядер дорсального таламуса амфибий с таковыми высших позвоночных не проведена, хотя, как полагают, часть nА, получающая ретинальный вход, может рассматриваться как гомолог дорсального ЛКТ млекопитающих.
Вентральный таламус амфибий содержит лежащее перивентрикулярно вентромедиальное ядро (VM) и расположенные латерально ядро Беллончи (nВ), вентролатеральное ядро (VL) и поверхностное желудочковое ядро, nucl. ventricular is superficialis (VS). Кроме того, в составе данной области описано несколько нейропилей, среди которых нейропиль Беллончи и нейропиль ЛКТ. Несмотря на наличие значительного ретинального входа nВ не образует проекций в теленцефалон, но направляет эфференты в тектум и заднюю таламическую зону, и на этом основании не может (вопреки ранее существовавшему мнению) рассматриваться как гомолог дорсального ЛКТ высших позвоночных. Дорсальная часть VL в прежних классификациях называлась круглым ядром и относилась к дорсальному таламусу. Однако ни распределение эфферентов, ни характер получаемых афферентов не доказывают гомологичности этой области одноименному ядру рептилий. Для ядер VM и вентральной части VL обнаружены афференты от области задвижки, а для последнего и от сетчатки. Эфференты обоих ядер направляются к тектуму, а ядра VM - и в спинной мозг. Поверхностное желудочковое ядро часто рассматривают как гомологичное вентральному ЛКТ млекопитающих из-за наличия ретинальных входов, однако существующих доказательств этого явно недостаточно. Некоторые авторы полагают, что вся медиальная часть вентрального таламуса амфибий может рассматриваться скорее как гомолог неопределенной зоны промежуточного мозга млекопитающих.
В вентральный таламус амфибий часто включают и задний
бугорок, представленный небольшой группой перивентрикулярно расположенных нейронов. Однако в отличие от вентрального таламуса эта область формирует проекции в теленцефалон, а в отличие от дорсального - проецируется на спинной мозг. Известные ее афференты происходят из медиального паллиума и стриатума. Такой характер связей не позволяет рассматривать задний бугорок как дериват дорсального или вентрального отделов, но вместе с тем значительно отличает его от одноименной части промежуточного мозга рыб, где основные связи приурочены к ниже расположенным отделам.
Строение таламуса высших позвоночных
Таламус рептилий весьма сложен и развитие структур его дорсального и вентрального отделов варьирует у представителей разных групп.
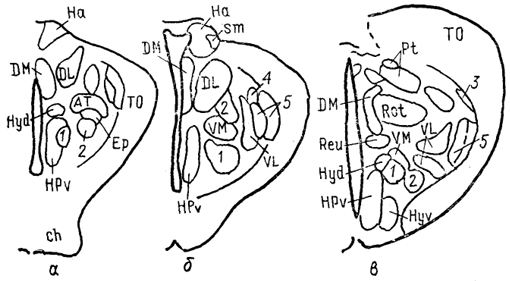
Рис. 70. Строение промежуточного мозга игуаны Iguana iguana (Butler, Northcutt, 1978).
1, 2 - пучки переднего мозга: медиальный (1) и латеральный (2); 3 - лентиформное ядро среднего мозга; 4, 5 - ЛКТ: дорсальное (4) и вентральное (5). а-в - срезы в рострокаудальном направлении.
Дорсальный таламус рептилий в ростральной части представлен дорсомедиальным и дорсолатеральным ядрами (DM и DL), в средней части располагаются круглое ядро, nucl. rotundus (Rot), и медиальное ядро, которое у многих смыкается по средней линии и составляет nucl. reuniens (Reu) (рис. 70). Каудальная часть дорсального таламуса включает медиальное заднее ядро (МР). Ядра DM и DL рассматриваются в качестве гомологов ядер передней группы таламуса млекопитающих на основании сходства их эфферентов, направляющихся в стриатум, дорсолатеральный паллиум и гиппокамп. Об их афферентах сведения немногочисленны, имеются указания на тектальные и соматические входы. Ядро Reu получает входы от торуса и представляет собой таламический отдел слуховой сенсорной системы
рептилий, его эфференты распределяются в медиальной части дорсального вентрикулярного края и в стриатуме (отметим, что проекции этого ядра исключительно ипсилатеральные). Круглое ядро - одно из наиболее крупных образований дорсального таламуса большинства рептилий (правда, у некоторых: змей оно мало или не описано вовсе). Входы к нему образованы тектумом, дорсолатеральным отделом дорсального вентрикулярного края и стриатумом. Эфференты адресованы латеральной части ДВК. Ядро МР является местом окончания спинно-таламических путей и проецируется на медиальную часть ДВК. (в этом усматривают сходство рептилий с птицами).
Вентральный таламус у рептилий включает в себя лежащую за преоптическим углублением area triangularis (AT), энтопедункулярное (Ер), вентролатеральное (VL) и вентромедиальное (VM) ядра, а также овальное ядро (Ov) и латеральное коленчатое тело (CGL). Последнее дифференцируют на дорсальную и вентральную части, каждая из которых получает ретинальные входы. Дорсальная часть обычно рассматривается как происходящая из дорсального таламуса, ее эфференты распределяются в дорсальной коре. Вентральная часть получает также тектальные афференты, а направление ее эфферентов неясно. В составе вентральной части выделяют медиальную клеточную пластинку и латерально расположенный нейропиль. Как тектальные, так и ретинальные афференты билатеральны.
Ядра VM и VL часто относят к структурам дорсального таламуса, поскольку (на основании соматосенсорных проекций от спинального лемниска) их гомологизируют с вентробазальньш комплексом млекопитающих. Вместе с тем к этим ядрам, а также к энтопедункулярному подходят проекции от стриатума (от мозжечка - для VL). У млекопитающих такой характер афферентов обнаруживается как в вентральном, так и в дорсальном таламусе, а различия касаются направленности эфферентов. В отношении рептилий сведений для окончательного отнесения ядер к тому или другому отделу недостаточно. Имеются данные о проекциях ядра VL в ростральные области паллиума. Помимо этого у некоторых рептилий оно получает вход сетчатки и проецируется в тектум, что позволило высказать мнение о гомологичности его неопределенной зоне. Ядро VM, как полагают, сходно с субталамическими полями Фореля. Однако обилие вариантов строения затрудняет решение вопросов гомологизации даже среди рептилий, а тем более между рептилиями и* птицами, недостаток же фактических данных, в частности сведений об эфферентах, не позволяет провести разделение этих образований в пределах таламуса.
В целом в таламусе рептилий возрастает число ядерных образований по сравнению с низшими позвоночными, хотя в структурном отношении сохраняются некоторые примитивные черты организации: существование нейропилей, ветвление дендритов
нейронов в пределах других ядер - вероятно-, древний способ обеспечения конвергенции разномодальной информации (рис. 71). Совершенно определенно в таламусе рептилий можно обнаружить выраженную направленность связей в теленцефалон, причем эти связи исключительно ипсилатеральны (в отличие от имеющихся даже у анура). Однако гомологизация структур таламуса рептилий с таламическими ядрами высших оказывается чрезвычайно трудной даже в отношении хорошо изученных отделов зрительной сенсорной системы - число ядер таламуса, получающих ретинальные входы, у рептилий оказалось весьма значительным, особенно у видов с развитым зрением.
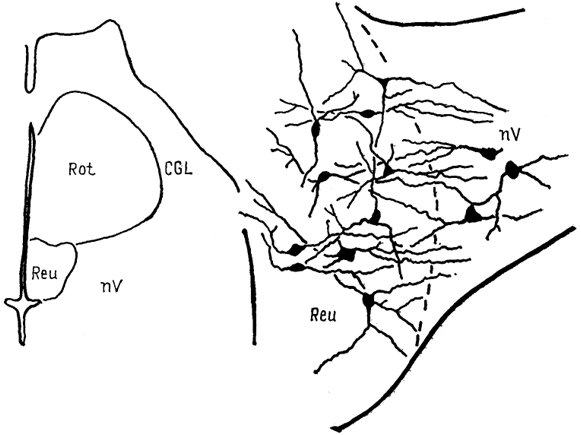
Рис. 71. Нейронные взаимоотношения в таламусе черепахи Emys orbicularis (Белехова, Туманова, 1988).
В пределах дорсального таламуса птиц выделяют дорсолатеральное, дорсомедиальное и дорсальное промежуточное ядра (DL, DM, DI), каждое из которых состоит из нескольких групп, обозначаемых как передняя, задняя, медиальная - DLA. DLP, DLM, DMA, DMP и др. и имеющих разную степень развития. Наиболее крупным компонентом таламуса птиц является круглое ядро (Rot). Под ним располагается nucl. triangularis (Tr), вентролатерально лежит прекомиссуральное ядро, nucl. pre-commissuralis principalis, а медиально-задневентральное ядро (VP). Достаточно велико, а у некоторых видов соразмерно с Rot овальное ядро, nucl. ovoidalis (Ov). Его развитие коррелирует со степенью выраженности среднемозгового слухового центра и с развитием слуховой системы. Латеральную часть дорсального таламуса составляет наружное мелкоклеточное ядро,
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|