- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 19 страница
В целом конечный мозг лучеперых рыб отличает высокий уровень структурно-функциональной организации. Он получает проекции различных сенсорных систем (обонятельной, зрительной, соматической), а через систему эфферентных связей принимает участие в регуляции многих физиологических реакций рыб. Вместе с тем, несмотря на попытки провести гомологизацию отделов конечного мозга лучеперых рыб и других позвоночных, задача эта однозначно не решена.
У амфибий в паллиуме выделяют медиальный, дорсальный и латеральный отделы. У бесхвостых амфибий паллиум отделяется от субпаллиума полосками белого вещества, у хвостатых и безногих эта граница выражена только в районе медиального паллиума.
У всех амфибий наиболее дифференцирован медиальный паллиум, где нейроны активно мигрируют в толщу стенки мозга, тогда как в других отделах они сосредоточены в основном в пе-ривентрикулярной зоне. У высших бесхвостых амфибий (жаб, жерлянок, квакш) в медиальном паллиуме цитоархитектонически выделяют две зоны: медиальную и дорсомедиальную.
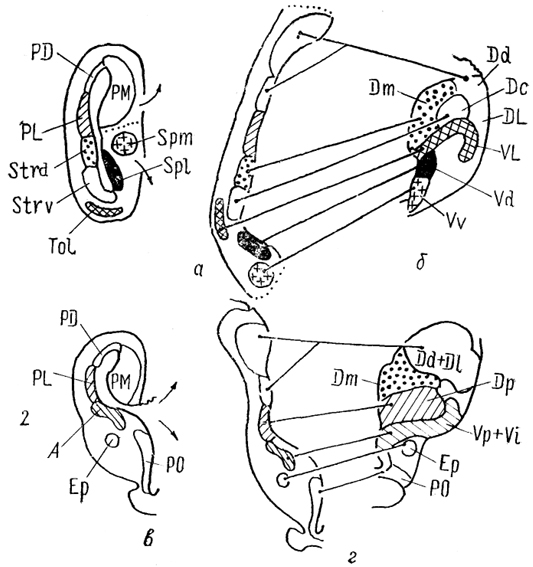
Рис. 86. Схема возможных гомологии структур инвертированного (а, в) и эвертированного (б, г) конечного мозга (Northcutt, 1981).
1 - ростральные и 2 - каудальные отделы полушарий.
Большинство данных, касающихся проводящих систем конечного мозга, получено на представителях бесхвостых амфибий, хвостатые и безногие в этом отношении менее изучены.
Афферентные проекции паллиума направляются главным образом в его медиальный отдел. По медиальному обонятельному тракту сюда поступает обонятельная информация, а по медиальному пучку переднего мозга через переднее ядро дорсального таламуса медиальный паллиум получает афференты зрительной, слуховой и соматосенсорной систем, а также гипоталамические проекции.
Эфференты медиального паллиума направляются в соседние зоны паллиума, септум, амигдалу (медиальную часть), вентральный таламус и гипоталамус и организованы в пучки, аналогичные своду, fornix, конечного мозга высших позвоночных. Отделы медиального паллиума полушарий соединены друг с другом паллиальной комиссурой - аналогом комиссуры гиппокампа высших позвоночных.
Дорсальный паллиум имеет обширные двусторонние связи с
соседними зонами паллиума, прямые обонятельные и таламические проекции отсутствуют. Латеральный паллиум является основной областью окончаний обонятельных проекций, достигающих его по латеральному обонятельному тракту. Эфференты широко распределяются среди паллиальных и субпаллиальных образований, а также направляются в ипсилатеральную обонятельную луковицу.
Субпаллиальные отделы конечного мозга амфибий включают стриатум, септум, обонятельный бугорок, амигдалу (медиальную часть), переднее обонятельное ядро и nucl. accumbens.
Стриатум - самый большой отдел субпаллиума амфибий - образован мелкими изодендритными нейронами, сосредоточенными в перивентрикулярной зоне. У бесхвостых амфибий стриатум разделяется на дорсальный и вентральный отделы, различающиеся плотностью расположения клеток. Основные проводящие пути стриатума проходят по системе латерального пучка переднего мозга. В таламусе источниками афферентных проекций в стриатум являются ядра центрального и латерального отделов. Одно из этих ядер (латеральное) связано с тектумом и передает в стриатум зрительную информацию. Незначительные афферентные проекции стриатум получает из вентрального гипоталамуса, преоптической области, латеральной части амигдалы и энтопедункулярного ядра. Особо следует отметить формирование у амфибий двусторонних связей стриатума с претектальной областью и тегментумом, что позволяет сравнивать их со стриато-нигро-стриатарными связями мозга высших позвоночных. С паллиумом стриатум амфибий не связан.
Септум располагается на медиовентральной стенке полушария и разделяется на два ядра: медиальное и латеральное. Степень их развития у представителей разных отрядов амфибий различна. У хвостатых (например, у Triturus vulgaris медиальное ядро развито слабо и практически не отделено от латерального, а нейроны обоих ядер относятся к разновидностям клеток изодендритного типа. У бесхвостых амфибий (Rana temporaria, Bufo bufo) медиальное ядро четко отделяется от латерального, а в их составе появляются многочисленные радиальные и биполярные нейроны (дендриты последних связывают два септальных ядра). По медиальному пучку переднего мозга в септум поступает соматосенсорная и зрительная информация из таламуса, а по медиальному обонятельному тракту - обонятельная (в ростральные отделы септума). Эфференты направляются в преоптическую область, вентральный таламус и гипоталамус. Септум имеет также обширные двусторонние связи с медиальным и дорсальным паллиумом.
Обонятельный бугорок достаточно подробно описан лишь в мозге бесхвостых амфибий, где он отчетливо выявляется в вентральных отделах полушария и представлен скоплением мелких слабодифференцированных нейронов. Основные афференты
поступают к нему по медиальному обонятельному тракту от ипси- и контрлатеральной обонятельной луковицы, а также от медиального паллиума. Эфференты практически не изучены; (они отмечены только в медиальном паллиуме). Этот отдел мозга амфибий часто сравнивают с разными структурами конечного мозга высших позвоночных (обонятельным бугорком, ядром диагональной связки Брока, частью медиального септального ядра).
Амигдала амфибий занимает в каудальных отделах полушария значительную часть вентролатеральной стенки мозга и разделяется иа два участка: медиальный (субпаллиальный) и латеральный (паллиальный). Медиальная амигдала имеет довольно сложный нейронный состав и формирует в основном двусторонние связи с медиальным паллиумом, стриатумом, гипоталамусом и таламусом. Латеральная амигдала представлена группой мелких веретеновидных нейронов, расположенных вблизи латерального желудочка, и является единственной структурой полушария, получающей проекции от добавочной обонятельной луковицы. Эфференты достигают гипоталамуса, вентрального таламуса, тектума.
Многие исследователи ранее рассматривали мозг современных амфибий как структуру, которая претерпела в процессе-эволюции определенное упрощение. Об этом свидетельствуют, в частности, перивентрикулярный характер расположения клеток в большинстве отделов паллиума и субпаллиума, отсутствие во взрослом состоянии корковой пластинки, большое количество изодендритных нейронов. Вместе с тем, в конечном мозге современных амфибий (особенно бесхвостых) есть признаки, говорящие о достаточно высоком уровне его структурно-функциональной организации. Так, основная и добавочная обонятельные луковицы имеют раздельные области представительства в полушариях (латеральный паллиум и латеральная часть амигдалы соответственно). Формируются две области конвергенции разно-модальной сенсорной информации, поступающей в полушарие - в медиальном паллиуме и стриатуме. Полагают, что у амфибий в медиальном паллиуме закладывается система связей, аналогичная таковой в гиппокампе мозга высших позвоночных и являющаяся важным компонентом лимбической системы. Стриатум участвует в формировании связей между основными уровнями экстрапирамидной системы - теленцефальным и среднемоз-говым. Все это позволяет конечному мозгу принимать участие в регуляции важнейших функций организма амфибий.
В целом для конечного мозга низших позвоночных характерны различные варианты организации: это и высокодифференцированный мозг некоторых хрящевых рыб, необычный по структуре мозг кистеперых рыб и эвертированный мозг лучеперых рыб, что свидетельствует о независимой и параллельной эволюции конечного мозга в разных группах анамний. При этом
базовой чертой организации их конечного мозга является наличие в его структурах зон представительства разных сенсорных систем (обонятельной, зрительной, слуховой, соматической) и, как следствие этого, выполнение конечным мозгом функций одного из интегрирующих центров головного мозга.
* Вентральная область полушарий конечного мозга лучеперых рыб не подвергается эверсии и ее положение сходно с таковым вентральной (субпаллиальной) области мозга инвертированного типа.
§ 2. КОНЕЧНЫЙ МОЗГ ВЫСШИХ ПОЗВОНОЧНЫХ
В трактовке морфофункциональной организации конечного мозга амниот, особенно рептилий и птиц, произошли значительные изменения. Ряд крупных образований, занимающих в полушарии субвентрикулярное положение и рассматриваемых ранее как субпаллиальные, на основании современных данных по эмбриогенезу, гистохимии, нейронной структуре и связям относят к отделам паллиума.
В конечном мозге рептилий паллиальными формациями являются собственно паллиальная (корковая) пластинка, разделенная на медиальный (гиппокампальный), дорсальный (общий) и латеральный (пириформиый) отделы, а также структуры так называемого дорсального вентрикулярного края (рис. 87).
Паллиум у разных рептилий имеет различную степень дифференцировки. У ящериц границы его отделов определяются достаточно четко по наличию зон перекрытия между медиальным и дорсальным и между дорсальным и латеральным паллиумом. У черепах, крокодилов и змей такого рода переходных зон нет и границы отделов паллиума определяются по изменению их внутренней структуры (цитоархитектонике, нейронному составу и связям).
В медиальном паллиуме большинства рептилий выделяют два участка: вентральный, мелкоклеточный (Ml), и дорсомедиальный, крупноклеточный (М2), составленные несколькими типами нейронов, в том числе пирамидоподобными. Источниками афферентов являются структуры конечного мозга (дорсальный и латеральный паллиум, ДВК, септум), переднее дорсолатеральное ядро таламуса, мамиллярные тела, ядра шва. Эфферентные проекции направляются в септум, амигдалу, передние отделы таламуса, гипоталамус. Через переднюю комиссуру медиальный паллиум связан с одноименной зоной паллиума другого полушария, а по системам внутриполушарных связей - с дорсальным паллиумом и ДВК.
Дорсальный паллиум рептилий разделяется на дорсомедиальную и дорсолатеральную части. Дорсомедиальная сходна у всех рептилий по структуре и связям, дорсолатеральная различна у чешуйчатых рептилий и черепах. У последних ее нейроны получают проекции по системе латерального пучка переднего мозга от дорсальной части ЛКТ и из вентрального таламуса, тогда как у ящериц в дорсолатеральный отдел дорсального паллиума
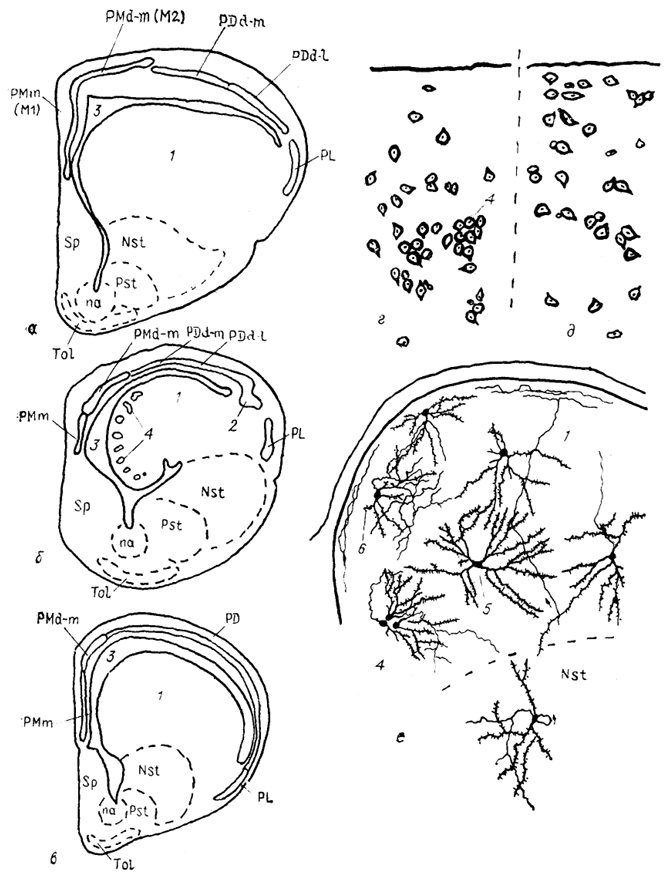
Тис. 87. Структура конечного мозга чешуйчатых рептилий (а), черепах (б) и крокодилов (в), цитоархитектоника дорсо-вентрикулярного края (ДВК) первого (г) и второго (д) типов и фрагмент нейронной структуры ДВК гадюки (е) (Обухов, Санькова, 1986; Ulinski, 1990).
1 - ДВК, 2 - паллиальное утолщение, 3 - латеральный мозговой желудочек, 4 - клеточные группы, 5 - радиальный густоветвистый и 6 - короткоаксонный нейроны ДВК.
по системе медиального пучка проецируется только дорсолатеральное ядро таламуса. Другие афференты дорсального паллиума приходят из ростральных отделов ДВК, паллиального утолщения, гипоталамуса, вентральной тегментальной области, ядер шва и голубого пятна.
Основная часть эфферентов направлена в гипоталамус, медиальный паллиум, септум. Отмечены проекции в стриатум, ДВК, амигдалу, переднее обонятельное ядро, основную и добавочную обонятельные луковицы, дорсальный таламус (дорсомедиальное и дорсолатеральное ядро). Комиссуральные связи обеспечивает передняя комиссура.
Самый латеральный отдел дорсального паллиума образует так называемое паллиальное утолщение специфического строения. Корковая пластинка утрачивает здесь слоистую структуру, а нейроны группируются в несколько крупных скоплений. Наиболее выражено паллиальное утолщение у черепах. В эту область проецируются волокна из дорсального таламуса. Так, через круглое ядро в паллиальное утолщение мозга ящериц поступает зрительная информация. Высказано предположение, что у рептилий в эволюции сформировалось несколько самостоятельных таламо-кортикальных путей в медиальный и дорсальный паллиум, паллиальное утолщение, ДВК.
Долгое время дорсальный паллиум (особенно его латеральные участки) рассматривали как возможный гомолог специфических сенсорных зон неокортекса млекопитающих. Однако система двусторонних связей с медиальным паллиумом и септумом, а также наличие обонятельных проекций из латерального паллиума, свидетельствуют о том, что этот отдел полушария (по крайней мере, у ящериц) наряду с медиальным паллиумом является теленцефалическим компонентом лимбической системы.
Латеральный паллиум рептилий занимает часть латеральной стенки полушария и представлен популяцией радиальных и горизонтальных нейронов изодендритного типа. Он имеет, как и другие отделы паллиума, отчетливое корковое строение. Латеральный паллиум получает прямые обонятельные проекции. Остальные афференты приходят из ростральных отделов ДВК, амигдалы и ядра латерального обонятельного тракта. Эфференты направляются в дорсальный паллиум и мелкоклеточную часть медиального паллиума, где оканчиваются на базальных дендритах ее нейронов, а также в амигдалу.
Дорсальный вентрикулярный край (ДВК) есть в полушариях конечного мозга всех рептилий и состоит из двух отделов: переднего и заднего. Передний ДВК относится к собственно паллиальным формациям, а задний, включающий такое крупное образование, как сферическое ядро, - к амигдалярному комплексу.
Размеры и структура переднего ДВК у рептилий, как из разных групп, так и у близкородственных видов, сильно варьирует.
Так, у ящериц Dracomorpha ДВК Занимает от 22 до 44%, объема всего полушария, а у ящериц Lacertomorpha - от 7 до 12%. Цитоархитектонически выделяют два типа ДВК. К первому типу относят ДВК, в составе которого выделяют поверхностную клеточную пластинку, образованную скоплением групп нейронов, и центральное ядро, имеющее диффузное строение (рис. 87). Такой тип ДВК отмечен у черепах, гаттерий, ряда змей и некоторых других чешуйчатых (сем. Gekkonidae, Lacertidae, Anguidae). В структуре ДВК второго типа отсутствует четкое разделение на клеточную пластинку и центральное ядро. Нейроны, как одиночные, так и в группах, равномерно распределены по всему ДВК. Этот тип ДВК встречается у крокодилов, некоторых змей и ящерицевых (сем. Agamidae, Iguanidae, Varanidae).
В ДВК рептилий обнаружены дискретные зоны проекций таламических ядер. Так, у безногой ящерицы желтопузика через, круглое ядро таламуса в латеральную зону ДВК поступает зрительная информация, а через nucl. reuniens в медиовентральную зону ДВК - соматическая и слуховая. На границе этих зон выявляется большое число полимодальных нейронов, что связано с явлением взаимопроникновения дендритов нейронов этих зон, где на них оканчиваются афференты другой модальности.
Подобные проекционные зоны обнаружены в мозге черепах и крокодилов, но, в отличие от ящериц, у них происходит обособление центральной зоны ДВК, получающей через медиальное заднее ядро таламуса соматическую информацию. Помимо проекций из таламуса, ДВК имеет обширные афферентные и эфферентные связи с другими отделами паллиума, стриатумом, амигдалой.
Таким образом, ДВК конечного мозга рептилий - высокодифференцированный отдел полушария, имеющий паллиальное происхождение. По наличию модальноспецифических сенсорных зон он сходен с неокортексом мозга млекопитающих, однако в отличие от ряда полей неокортекса, ДВК рептилий не имеет прямых проекций в моторные центры ствола и спинного мозга. Исследователи считают ДВК структурой, специфичной для конечного мозга рептилий, эволюция которой проходила независимо и параллельно развитию других паллиальных формаций мозга высших позвоночных.
К субпаллиуму конечного мозга рептилий относят стриатум и ряд структур вентромедиальной стенки полушария: обонятельный бугорок, септум, nucl. accumbens, и амигдалу, ядра которой расположены в вентролатеральной зоне полушария.
Стриатум разделяется на два отдела: дорсальный, получивший название неостриатум, neostriatum, и вентральный - палеостриатум, paleostriatum (рис. 87). Неостриатум составляет незначительную часть стриатарного комплекса и хорошо выражен у всех рептилий. Размеры и топография неостриатума подвержены довольно слабым межвидовым колебаниям. В его составе:
не выделяется каких-либо отдельных образований, а по уровню нейронной дифференцировки он значительно уступает соседнему ДВК. Палеостриатум всех изученных рептилий разделяется на две части: медиальную (мелкоклеточную) и латеральную (крупноклеточную).
Среди проекций неостриатума наиболее важными являются двусторонние связи с дорсальным паллиумом и передним ДВК, аналогичные неостриато-корково-неостриатарным связям конечного мозга млекопитающих. Обнаружены также двусторонние связи стриатума с тегментумом среднего мозга (ядром ТРС)Г которые сравниваются со стриато-нигро-стриатарными связями млекопитающих. Кроме того, показано, что проекции сенсорных, таламических ядер, проходящие в паллиум и ДВК, дают в стриатуме коллатерали (рис. 88). Основная масса эфферентных волокон неостриатума проходит в палеостриатум, который в свою очередь образует главные эфферентные тракты стриатума. Так, у ящерицы Gekko gekko были отмечены проекции в передние и задние энтопедункулярные ядра, в вентромедиальное и вентро-латеральное ядра таламуса. У черепах и крокодилов (кайманы) палеостриатум связан с ядром задней комиссуры, которое проецируется в тектум. Эфферентные проекции палеостриатума рептилий прослеживаются также в такие структуры, как ЦСВ среднего мозга, ядра шва, интерколликулярное ядро, латеральное ядро мозжечка и ретикулярную формацию.
Таким образом, структура и система связей стриатума позволяют рассматривать его как сложный корреляционный центр полушарий рептилий. Особо следует отметить формирование впервые у позвоночных системы двусторонних стриато-паллиальных связей.
Септум мозга разных рептилий имеет единый план строения, сходную нейронную организацию и состоит из медиального и латерального ядер. В септуме описано два основных типа нейронов: мультиполярные редковетвистые и веретеновидные биполярные. Главными проекциями септума являются билатеральные двусторонние связи с медиальным паллиумом и контрлатеральные афференты из дорсального паллиума, обонятельного бугорка и амигдалы. По медиальному обонятельному тракту ипсилатерально в септум приходят волокна из обонятельной луковицы. В составе медиального пучка переднего мозга аксоны нейронов септума достигают дорсального таламуса, преоптической области, гипоталамуса. Таким образом, септум является важным связующим звеном между паллиальными формациями и диенцефалоном и входит вместе с медиальным паллиумом в группу лимбических структур мозга.
Вентральная область полушария конечного мозга рептилий представлена несколькими мелкими ядрами, среди которых наиболее полно изучены обонятельный бугорок и nucl. accumbens.
Обонятельный бугорок рептилий получает афферентные проекции

Рис. 88. Основные связи стрио-паллидарной системы мозга рептилий (а) и птиц (б) (Белехова, 1986).
1 - дорсо-вентрикулярный край, ALa, ALp - ядра лентикулярной петли, DcP – ядро задней комнссуры, DIP - заднее промежуточное ядро таламуса, SpL -спириформное ядро претектальной области.
из обонятельной луковицы (основной), переднего обонятельного ядра, латерального и медиального паллиума. Эфференты бугорка направляются в медиальный паллиум и таламус.
Nucleus accumbens рептилий отличается простотой цитоархи-тектонической и нейронной организации, будучи представлен небольшой группой мелких изодендритных нейронов. Основные афференты он получает от септума, обонятельного бугорка, ДВК, дорсолатерального таламуса, а его эфференты направляются в вентральные районы палеостриатума, преоптическую область, латеральный гипоталамус и вентральную тегментальную область.
Амигдала рептилий представляет собой сложный комплекс ядер, связи и происхождение которых во многом неясны. У рептилий она расположена в вентролатеральной части полушария в задней трети конечного мозга. Выделяется от двух до семи
ядер, которые могут объединяться в центральный и базальный комплексы. В состав амигдалы мозга рептилий включают и так называемое сферическое ядро - задний отдел ДВК (оно есть не у всех рептилий и наиболее отчетливо выражено у змей и ящериц-макросматиков). Сферическое ядро получает волокна добавочной обонятельной луковицы и вовлечено в интеграцию обонятельной и соматической информации. Описаны эфферентные связи ядра с преоптической областью и каудальным гипоталамусом. У рептилий помимо филогенетически древних ядер амигдалы, связанных с обонятельной системой (в их число попадает и сферическое ядро), есть новые ядра, связанные с ДВК и корковыми формациями.
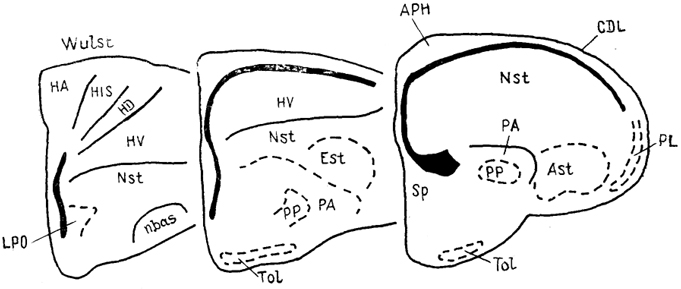
Рис. 89. Строение полушарий мозга голубя Columba livia на серии фронтальных срезов (Reiner, 1985).
Конечный мозг птиц имеет необычное по сравнению с мозгом других высших позвоночных строение. Латеральные желудочки развиты слабо, почти все полушарие занято диффузными клеточными массами, среди которых выделяют такие структуры, как гипер-, нео-, архи- и палеостриатум (рис. 89). Они разграничены узкими полосками белого вещества.
К собственно паллиальным формациям относят кортикоидную пластинку, расположенную на дорсомедиальной поверхности полушария, и гипер- и неостриатум, которые ранее считали субпаллиальными структурами.
Кортикоидная пластинка мозга птиц максимально развита в каудальных отделах полушария и состоит из дорсомедиального, дорсолатерального отделов и участка латерального паллиума. У многих птиц кортикоидная пластинка имеет сложный нейронный состав и развитую систему связей. Здесь обнаружены пирамидоподобные нейроны и шипиковые и бесшипиковые коротко-аксонные клетки идиодендритного типа. Дорсомедиальный отдел получает афференты от гиперстриатума, дорсолатерального отдела паллиума, архистриатума, гипоталамуса, ряда ядер ствола
мозга (ядро шва, голубое пятно) и одноименной структуры из соседнего полушария. Эфференты направляются в септум, дорсолатеральный паллиум, дорсальные отделы гипер- и архистриатума.
Все это дает основание проводить гомологию дорсомедиального отдела паллиума конечного мозга птиц с медиальным паллиумом мозга рептилий и других позвоночных. Структура и связи дорсолатерального отдела кортикоидной пластинки изучены недостаточно.
Латеральный паллиум птиц развит слабо, однако обнаруженные здесь проекции обонятельной луковицы позволяют сравнивать его с латеральным паллиумом рептилий.
Цитоархитектоническая структура гипер- и неостриатума достаточно сложна. Существуют две основные классификации структур полушарий мозга птиц, согласно которым в нем выделяют ряд отделов или полей, степень развития которых сильно варьирует у разных птиц.
Hyperstriatum accessorium (классификация С. Херрика), или поле В (классификация М. Розе) - самый дорсальный отдел гиперстриатума. У большинства изученных видов он разделяется на две части: дорсальную, имеющую кортикоидную структуру и тесно связанную с кортикоидной пластинкой, и вентральную - ядерного типа. Обе части содержат как одиночные нейроны, так и клеточные скопления разного типа. Наиболее значительна дорсальная часть гиперстриатума у пингвинов. Hyperstriatum intercalatus superior (поле А) - наиболее вариабельный по размеру и степени дифференцировки отдел. У одних видов (голуби) он развит слабо, у других (совы, пингвины, вороны) четко выражен. Hyperstriatum dorsale (поле С) есть у всех птиц и отличается наличием большого количества крупных нейронов и нейронных комплексов. Hyperstriatum ventrale (поле D) - самое крупное поле гиперстриатума по размеру как самой структуры, так и составляющих его элементов. Neostriatum (поле G) - структура, простирающаяся до самых каудальных отделов полушария. В его составе выделяют несколько участков: в медиальной части каудального отдела - поле L; в средних отделах - эктостриатум, ectostriatum (поле Е). От соседних участков неостриатума последний отличается однообразием клеточных форм и разделяется на центральную и периферическую части.
Ростральные отделы гиперстриатума образуют особую структуру - дорсальное возвышение, Wulst. По степени его развития и положению выделяется два типа полушарий конечного мозга птиц (рис. 90). Первый характеризуется гипертрофией Wulst и смещением его в ростральные отделы полушария, второй - более слабым развитием и сдвигом в теменно-затылочную область. По первому типу построен конечный мозг голубиных, пингвинов, куриных, чайковых, птиц, дневных хищников, совиных,
воробьиных. Интересно отметить, что у перечисленных групп птиц кортикоидная пластинка паллиума развита по-разному. Так, у голубиных, куриных, пингвинов она выражена относительно хорошо, а у воробьиных (особенно у врановых) - плохо. По второму типу построен мозг представителей отрядов ржанковых, пастушковых, голенастых, веслоногих, пластинчатоклювых, а также у дятлов и попугаев. У этих птиц хорошо развита кортикоидная пластинка.
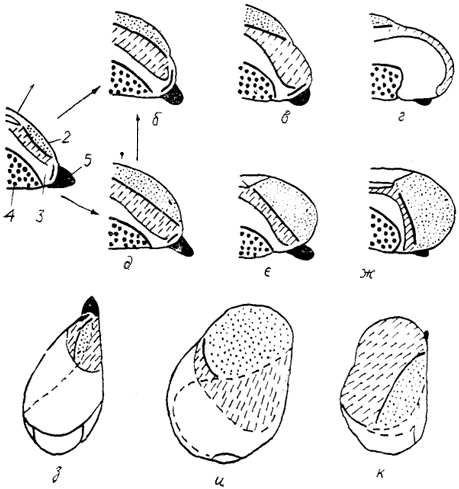
Рис. 90. Два типа организации конечного мозга птиц (Stingelin, 1958).
а - исходный (гипотетический) тип строения полушарий; б-г - полушария второго типа у морского зуйка (б), белого аиста (в) и бекаса (г); д-ж - полушария первого типа у голубя (д), козодоя (е) и грача (ж); з-к - вид мозга сверху: исходный тип (з), у грача (и) и бекаса (к). 1-5 - отделы полушария: 1 - Wulst, 2 - вентральный гиперстриатум, 3 - неостриатум, 4 - палеостриатум, 5 - обонятельная луковица.
Относительный объем полей гиперстриатума сильно варьирует как у представителей разных отрядов, так и в пределах одного отряда или семейства.
Функциональная организация гипер- и неостриатума мозга птиц также очень сложна. В этих структурах обнаружены двойные зоны представительства ряда сенсорных систем. Зрительная информация достигает ростральных отделов НА через дорсолатеральное переднее ядро таламуса, а через круглое ядро - центральной части эктостриатума. Соматосенсорная информация поступает в ростральные отделы НА через дорсальное промежуточное ядро таламуса, а через мультисенсорное дорсолатеральное заднеталамическое ядро - в район на границе каудального
неостриатума и вентрального гиперстриатума. Эти две зоны значительно отличаются друг от друга. Нейроны сомато-сенсорной зоны в ростральном гиперстриатуме имеют относительно небольшие рецептивные поля. При этом ростральные отделы тела животного представлены в дорсальной части зоны, каудальные - в вентральной. Нейроны второй соматосенсорной зоны имеют обширные рецептивные поля, получая соматическую информацию от половины, а иногда и всего тела, и являются при этом полимодальными, воспринимая поступающую в соседние районы полушария слуховую и зрительную информацию. Морфологически это обеспечивается длинными дендритами нейронов, выходящими далеко за пределы соматосенсорной зоны. Обе зоны связаны друг с другом двусторонними связями.
В каудальном неостриатуме (поле L) обнаружены входы; слуховой сенсорной системы, волокна которой через овальное ядро таламуса проецируются также в архистриатум, а через медиальную часть дорсолатерального ядра - в ростральные отделы несстриатума.
В свою очередь Wulst и другие поля гипер- и неостриатума образуют обширные связи как внутри полушария, так и с другими отделами мозга. В частности, зрительные зоны Wulst образуют афференты в эктостриатум (поверхностную зону), палеостриатум, дорсальный и вентральный таламус, соседние поля гиперстриатума, претектальную область, тектум. Обнаружены прямые проекции из гиперстриатума в моторные ядра ствола и спинной мозг, аналогичные кортико-спинальным проекциям мозга млекопитающих.
Степень развития полей гиперстриатума (особенно района Wulst) и его нейронных комплексов коррелирует с уровнем сложности поведения птиц. При этом кортикоидная пластинка, гипер- и неостриатум, являясь паллиальными по происхождению, различаются не только по структуре, но и по участию в организации разных поведенческих реакций. Это свидетельствует о высоком уровне дифференцировки паллиальных структур конечного мозга птиц.
У птиц, как и у рептилий, к собственно стриатарным структурам относят палеостриатум и образования вентромедиальной стенки полушария (обонятельный бугорок, септум, nucl. accumbens, часть амигдалы).
В составе палеостриатума различают два поля: палеостриатум возрастающий, paleostriatum augmentatum (поле Н) и палеостриатум примитивный, paleostriatum primitivum (поле J) (рис. 89). Поле РА четко отделяется от вышележащего неостриатума полоской белого вещества и состоит из мелких и средних нейронов, как одиночных, так и образующих скопления (в их состав входит 3-6 нейронов). Поле РР имеет нечеткие границы и находится как бы внутри поля РА. Нейронный состав палеостриатума птиц сходен с таковым у рептилий.
Афферентные проекции поле РА получает из зоны РР, неостриатума, гиперстриатума, таламуса, среднего мозга. Многие из этих связей двусторонние. Основные эфферентные связи стри-атума образованы структурами поля PP. Так, обнаружены связи РР с ядром SPL претектальной области, которое связано с тектумом. Таким образом осуществляется связь с моторной системой мозга птиц, поскольку нейроны тектума образуют нисходящие проекции к моторным центрам ствола и спинного мозга.
Стриато-таламо-стриатарные связи осуществляются у птиц через заднее промежуточное ядро, которое получает мозжечковые афференты и проецируется в гиперстриатум, а также через ядра лентикулярной петли. Сформированы двусторонние связи РА и РР с ядром ТРС тегментума среднего мозга, которые гомологичны стриато-нигро-стриатарным связям конечного мозга рептилий и млекопитающих (рис. 88).
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|