- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 18 страница
произошла редукция теменного органа, а пинеальный, утрачивая светочувствительные элементы, превратился в железу внутренней секреции - эпифиз.
Отличительной особенностью этого отдела промежуточного мозга является его асимметрия, которую ранее пытались объяснить ранней атрофией левого парапинеального органа, однако сейчас показано, что у представителей разных групп позвоночных асимметрия комплекса разная: правосторонняя гипертрофия у круглоротых и левосторонняя - у хрящевых рыб и амфибий.
У круглоротых пинеальный и парапинеальный органы выражены различно. Пинеальный хорошо развит у миног, а у Мordacia mordax, которую считают промежуточной формой между миногами и миксинами,- отсутствует. В составе поводковых ядер выделяют перивентрикулярный и поверхностный отделы. Последний справа значительно превосходит по размерам соответствующий отдел в левых поводковых ядрах, что и определяет асимметрию комплекса. Афференты, подходящие к поводкам, идут в составе так называемых медуллярных полосок, striae medullares, и происходят главным образом из конечного мозга и частично из гипоталамуса. Эфференты формируют tr. habenulo-interpeduncularis, адресованный интерпедункулярному ядру "среднего мозга. Этот путь называют также отогнутым пучком Мейнерта, fasc. retroflexus Meynerti. В его составе имеются миелинизированные и немиелинизированные волокна.
У хрящевых рыб (например, у акулы Scyliorhinus caniculaпинеальный орган представлен длинной трубкой с дистальным закрытым отделом, лежащим на поверхности черепной коробки. Ее проксимальный конец открыт в полость III желудочка в районе между поводками и задней комиссурой. Паренхима пи-неального органа содержит фоторецепторы, опорные и ганглионарные клетки. Последние образуют пинеальный тракт, tr. pinealis, который идет к задней комиссуре и частично к поводковому комплексу. Точное место окончаний волокон не определено. Поводковые ядра составляют у хрящевых рыб ростральный отдел эпиталамуса, левые ядра комплекса превосходят правые. Характерное для большинства позвоночных разделение на медиальное и латеральное ядра у акул лишь намечается. Обе части комплекса соединяются по средней линии поводковой комиссурой, в составе которой у некоторых акул располагаются нейроны, подобные клеткам собственно ядер поводков и составляющие ядро Беллончи. У скатов оно отсутствует. Основные афференты приходят из конечного мозга (по мозговым полоскам), тектума и тегментума (текто- и тегменто-хабенулярные тракты).
У амфибий и лучеперых рыб эпиталамус состоит из комплекса дорсального и вентрального хабенулярных ядер, причем у амфибий, как отмечалось выше, наблюдается левосторонняя
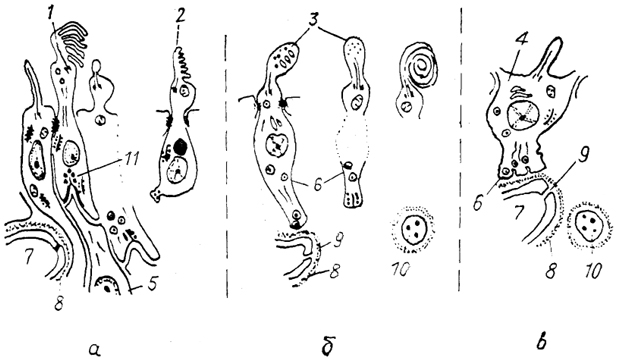
Рис. 81. Схема организации пинеалоцитов у анамний (а), зауропсид (б) млекопитающих (в) (Oksche, 1988).
1 - типичный фоторецепторный пинеалоцит, 2 - серотонинсодержащий пинеалоцит, 3 - модифицированный пинеалоцит, 4 - пинеалоцит млекопитающих, 5 - нейрон, 6 - секреторные гранулы, 7 - капилляр, 8 - базальная мембрана, 9 - поры в эндотелии, 10 - симпатическое нервное окончание, 11 - специализированный синапс.
асимметрия. Эфференты поводковых ядер направляются к интерпедункулярному ядру, а часть их проходит его без переключений и адресуется глубокому и дорсальному ядрам покрышки среднего мозга.
У высших позвоночных ядра поводков выражены не столь четко, как у низших, но характер их связей остается прежним. Афференты в основном происходят из теленцефальных образований, а эфференты распределяются среди структур среднего мозга: в интерпедункулярном ядре, ретикулярной формации, а также (у млекопитающих - от латерального ядра) в области мезенцефалического ядра шва.
У высших позвоночных прослеживаются регрессивные изменения в структуре эпифиза. Они выявляются в пределах каждого класса и особенно среди млекопитающих. Их эпифиз прикрепляется к поводковому комплексу и содержит клетки, получившие наименование пинеалоциты, а также интерстициальные клетки, подобные глиальным. Полагают, что пинеалоциты являются дериватами сенсорных клеток (рис. 81), которые опосредовали воздействие световых раздражителей на эндокринную систему, и таким образом являлись фотонейро-эндокринными элементами.
Смена функций, приведшая к превращению пинеального органа в железистое образование - эпифиз, определила значительные изменения и в структуре основных клеток. У птиц и млекопитающих
наиболее значительные изменения произошли в наружных сегментах пинеалоцитов. В результате у птиц и млекопитающих эти клетки, как полагали, утратили способность к реакции на свет. Но в последнее время показано, что пинеалоциты птиц сохранили способность к синтезу специфических фоторецепторных белков и реакции на свет. Пинеалоциты птиц и млекопитающих - типичные секреторные железистые элементы, синтезирующие мелатонин. Окончательным доказательством единства происхождения фоторецепторных пинеалоцитов низших позвоночных и секреторных клеток эпифиза высших позвоночных послужили иммуногистохимические данные о присутствии в типичных фоторецепторных пинелоцитах костистых рыб серотонина - предшественника мелатонина - гормона эпифиза.
Пока неясен механизм влияния секреторных клеток эпифиза птиц и млекопитающих на циркадную ритмику физиологических процессов, поскольку они непосредственно не подвергаются: воздействию световых раздражителей, но, возможно, это опосредуется через взаимодействие с ядрами гипоталамуса, получающими ретинальные входы (например, супрахиазматическое ядро). Нарушение этих связей приводит к нарушению циркадной; ритмики в работе эндокринных систем, связанных с эпифизом.
В целом структуры эпиталамуса в достаточно дифференцированном виде представлены уже у самых примитивных из ныне живущих позвоночных и на протяжении эволюции характеризуются постоянством состава и выполняемых функций. Так, пинеальный и парапинеальный органы, содержащие у низших, экстраретинальные фоторецепторы, обеспечивают контроль циркадной ритмики многих физиологических процессов. Эпифиз млекопитающих и птиц, лишенный рецепторных клеток, за счет гипоталамических входов продолжает выполнять роль, свойственную этому отделу мозга. Ядра поводков у всех позвоночных животных дают начало хабенуло-интерпедункулярной системе волокон. Последняя рассматривается как экстрагипоталамическая система пищевого поведения, а у высших как существенное, звено лимбической системы.
ГЛАВА 8
КОНЕЧНЫЙ МОЗГ
Конечный мозг, Telencephalon, анатомически разделяется на две части: каудальную (непарную), telencephalon impar, непосредственно переходящую в структуры промежуточного мозга, и ростральную, образованную парными полушариями, hemis-pherae telencephali. Все отделы конечного мозга развиваются в юнтогенезе как производные крыловидной пластинки. При этом уже на ранних стадиях формирования переднемозгового пузыря, матричная зона, где сосредоточены камбиальные элементы и откуда начинается активная миграция нейробластов, образующих :в дальнейшем те или иные структуры полушария, разделяется на две зоны: дорсальную и вентральную. Производными дорсальной матричной зоны являются формации паллиума, вентральной - субпаллиума.
В разных группах позвоночных процессы развития паллиальных формаций могут протекать по-разному, в результате чего образуется звертированный или инвертированный тип полушарий и конечного мозга в целом (см. рис. 4). Эвертированный тип полушарий встречается только у лучеперых рыб, а у остальных позвоночных они инвертированы. Субпаллиум процессу эверсии ни у кого из позвоночных не подвергается.
В ростральных отделах полушарий происходит образование ларных выпячиваний стенки мозга и формирование обонятельных луковиц.
Обонятельные луковицы, bulbus olfactorius, есть у большинства позвоночных (случаи их редукции у некоторых позвоночных, например ряда птиц и китообразных, рассматриваются эволюционно как вторичные). У круглоротых, большинства костистых рыб, амфибий, черепах, птиц и многих млекопитающих обонятельные луковицы непосредственно примыкают к основной части полушарий, тогда как у хрящевых, двоякодышащих, некоторых костистых рыб, ряда амфибий, большинства рептилий и у приматов они отнесены от нее на некоторое расстояние с помощью так называемых обонятельных ножек, по которым
проходят обонятельные тракты. Обычно обонятельные луковицы расположены относительно основной части полушарий рострально или ростро-вентрально, и только у акул лежат латерально (см. рис. 5). Их форма варьирует от овальной до конусовидной.
Обонятельная луковица у всех позвоночных имеет сходный план строения и состоит из нескольких концентрически расположенных клеточных и волокнистых слоев, структура которых значительно варьирует (рис. 82).
Волокна обонятельного нерва, войдя в луковицу, разветвляются и участвуют в формировании особых структур - клубочков, или гломерул, где они контактируют с дендритами различных нейронов луковицы. У низших позвоночных каждое волокно может участвовать в образовании нескольких клубочков, которые располагаются на разной глубине. В связи с этим клубочковый слой выражен неотчетливо. У высших позвоночных каждое волокно обонятельного нерва образует только одну гломерулу, а их слой компактный и ясно выражен.
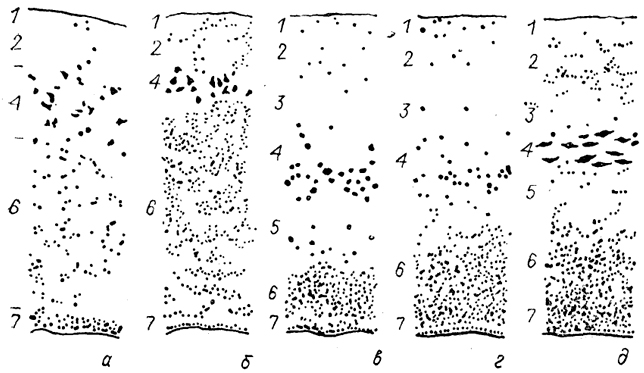
Рис. 82. Цитоархитектоника обонятельной луковицы миноги (а), акулы (б), двоякодышащей рыбы (в), хвостатой амфибии (г) и ящерицы (д) (Nieuwenhuys., 1967).
1-7 - слои обонятельной луковицы: обонятельных волокон (1), клубочковый (2), наружный волокнистый (3), митральный (4), внутренний волокнистый (5), зернистый (6) и эпендимный (7).
У большинства позвоночных в клубочковом слое присутствуют особые тормозные нейроны - межклубочковые. Их дендриты формируют в клубочках дендро-дендритные синапсы с отростками митральных нейронов, а также контактируют с обонятельными волокнами. Аксоны этих клеток направляются в соседние клубочки, где образуют тормозные синапсы.
Наиболее характерным слоем обонятельной луковицы является митральный, где сосредоточены специализированные митральные нейроны. Размеры, количество, характер их распределения и соответственно степень выраженности митрального слоя сильно варьируют у позвоночных. У одних (двоякодышащие рыбы, рептилии, птицы, млекопитающие) формируется четкий слой, у других (круглоротые, хрящевые и лучеперые рыбы, амфибии) митральные клетки расположены более или менее диффузно (рис. 82). У наземных позвоночных только один из дендритов каждого митрального нейрона входит в состав одного клубочка, тогда как у остальных - несколько дендритов принимают участие в формировании клубочков. Митральные нейроны из разных участков обонятельной луковицы связаны, по-видимому, с переработкой различной обонятельной информации. Аксоны митральных нейронов направляются в полушария.
Значительные изменения отмечены в организации зернистых, нейронов, формирующих у всех позвоночных гранулярный слой луковицы. У низших позвоночных они вместе с митральными нейронами участвуют в формировании клубочков, причем дендриты одной зернистой клетки входят в состав нескольких таких комплексов. У высших позвоночных зернистые клетки утрачивают связь как с клубочками, так и с полушарием (у них нет аксона), превращаясь таким образом в локальные интернейроны обонятельной луковицы. Дендриты зернистых клеток образуют вне зоны клубочков дендро-дендритные тормозные синапсы с митральными нейронами.
Для структуры обонятельной луковицы высших позвоночных, особенно млекопитающих, характерны особые нейроны - кисточковые. Они располагаются в наружном волокнистом слое и, подобно митральным нейронам, посылают один из дендритов в клубочек.
Эфферентные проекции обонятельной луковицы низших позвоночных сформированы аксонами митральных и зернистых нейронов, высших позвоночных - митральных и кисточковых Морфологически они оформлены в три обонятельных тракта: медиальный, латеральный и промежуточный. Последний выражен у высших позвоночных, состоит из аксонов в основном кисточковых нейронов и является комиссуральным. У других позвоночных комиссуральные связи осуществляются аксонами зернистых нейронов и проходят в составе передней комиссуры конечного мозга (у хрящевых рыб есть самостоятельный комиссуральный тракт, сходный с промежуточным обонятельным трактом высших позвоночных). Медиальный и латеральный тракты, топографически обособленные друг от друга, начинаются от разных групп митральных нейронов и проецируются в разные отделы головного мозга. У низших и высших позвоночных есть определенные различия в характере распределения окончаний обонятельных трактов (они рассматриваются в соответствующих
разделах главы). Здесь необходимо только подчеркнуть, что зона окончаний обонятельных проекций в структурах конечного мозга современных позвоночных ограничена.
Афферентная центрифугальная иннервация обонятельной луковицы обнаружена у всех позвоночных. В ее осуществлении принимают участие многие отделы конечного мозга (переднее обонятельное ядро, латеральный паллиум, обонятельный бугорок, ядра амигдалы), промежуточного мозга (гипоталамус, субталамус) и ряд ядер ствола (ядра шва, голубое пятно). В большинстве этих зон оканчиваются эфференты обонятельной луковицы. Центрифугальные волокна контактируют с дендритами зернистых нейронов.
Помимо основной обонятельной луковицы у ряда позвоночных (безногие и бесхвостые амфибии, многие рептилии, млекопитающие) есть дополнительная, bulbus olfactorius accessorius, которая по структуре сходна с основной и получает проекции от вомеро-назального органа (органа Якобсона). Необходимо отметить, что в дополнительной луковице даже у млекопитающих митральные нейроны располагаются диффузно, а их дендриты участвуют в формировании нескольких клубочков. Она имеет раздельные с основной луковицей зоны проекций в структурах мозга (в основном в амигдалярном комплексе), что определяет ее роль в организме.
Таким образом, обонятельные луковицы сохраняют в филогенезе позвоночных единый план строения, а их развитие идет по пути дифференциации нейронных элементов, все большего разграничения слоев и усложнения внутренних и внешних связей.
§ 1. КОНЕЧНЫЙ МОЗГ НИЗШИХ ПОЗВОНОЧНЫХ
У круглоротых морфологически выражены все основные отделы конечного мозга. В паллиуме полушария у миног описывают четыре отдела: медиальный, субмедиальный (субгиппокампальная доля), дорсальный и латеральный. Субпаллиум, представленный структурами переднего обонятельного ядра, стриатума и септума, у миног дифференцирован слабо. В срединной части каудального отдела полушария расположено преоптическое ядро (рис. 83).
Наиболее дифференцированным отделом полушария миног является медиальный, клетки которого имеют развитую систему дендритных ветвлений и образуют слоистую структуру. Субмедиальный, дорсальный и латеральный отделы паллиума значительно уступают по уровню нейронной дифференцировки медиальному и состоят из мелких нейронов изодендритного типа. Основная масса клеток сосредоточена вблизи латерального желудочка.
У миксин конечный мозг значительно отличается от такового миног. Это относительно крупный отдел, в котором полушария
трудно отделить от каудальной части конечного мозга из-за редукции, по-видимому, вторичной, латеральных желудочков. В полушариях выделяют лежащий медио-дорсально медиальный паллиум и латеральную часть, которую трудно сравнивать с какими-либо конкретными структурами паллиума мозга миног из-за необычного строения (в ней выделяют до шести слоев нейронов и их отростков). Субпаллиум миксин занимает в
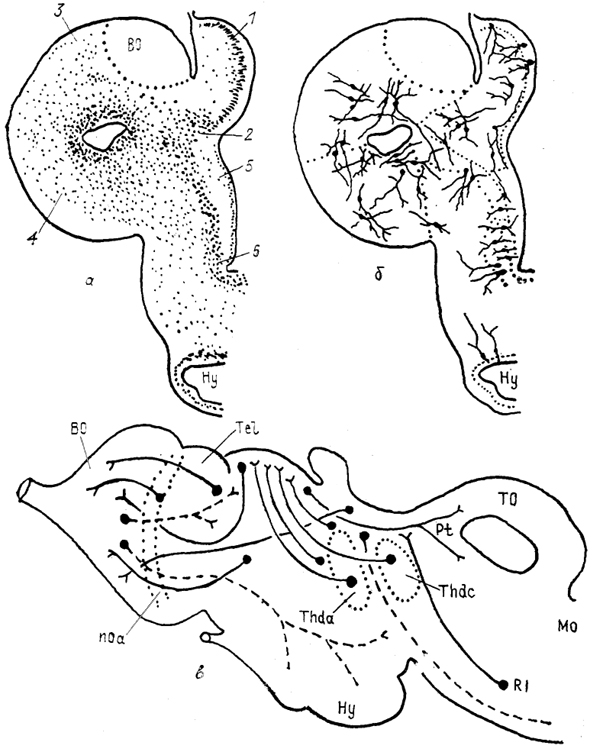
Рис. 83. Схемы строения (а), нейронной организации (б) и проводящих путей (в) конечного мозга миноги Lampetra fluviatilis (Haier, 1947; Поленова, 1990).
1 - примордиальный гиппокамп, 2 - субгиппокампальная доля, 3 - дорсальный и 4 - латеральный паллиум, 5 - стриатум, 6 - преоптическая зона.
полушарии медиовентральное положение и по структуре сходен с субпаллиумом миног. Система афферентных связей конечного мозга круглоротых представлена в основном обонятельными трактами, которые проецируются у миног билатерально в дорсальный и латеральный паллиум, а также контрлатерально в переднее обонятельное ядро и преоптическую область. Субгиппокампальная доля, вероятно, не получает прямых обонятельных проекций. Другие афференты конечного мозга происходят
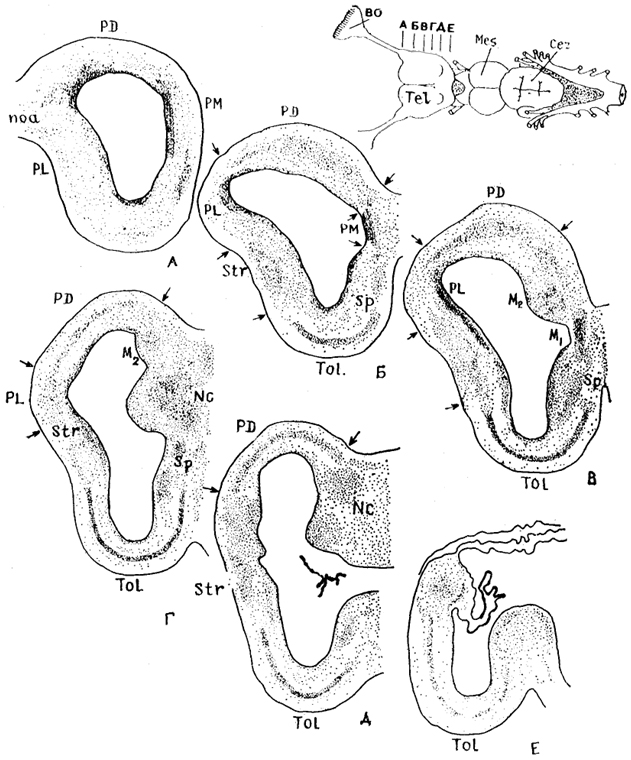
Рис. 84. Строение конечного мозга акулы Squalus acanthias (Обухов, 1987).
А-Е - уровни срезов мозга; Ml - медиальный и М2 - дорсомедиальный участки медиального паллиума, NC - центральные ядра. Стрелки - границы зон мозга.
из ядер дорсального таламуса и проводят информацию от зрительной и соматосенсорной систем (рис. 83). Они оканчиваются в областях полушария, как получающих обонятельные входы, так и свободных от них.
Эфференты паллиума осуществляют центрифугальную иннервацию обонятельной луковицы, кроме того, медиальный паллкум связан с преоптической областью, гипоталамусом и претектальной областью среднего мозга.
Таким образом, у круглоротых сформированы все основные отделы конечного мозга, характерные для других позвоночных, которые получают проекции от структур обонятельной, зрительной, ссматосенсорной систем. Медиальный отдел паллиума мозга круглоротых сравнивается с медиальным (гиппокампальным) паллиумом мозга челюстноротых позвоночных, субгиппокампальный с дорсальным, а дорсальный и латеральный отделы - с латеральным пириформным паллиумом. Вместе с тем современные круглоротые являются продуктом длительной эволюции, проходившей во многом независимо и параллельно эволюции в линии челюстноротых позвоночных, что не позволяет считать их мозг подобным мозгу предков позвоночных.
В обширном надклассе Рыбы встречаются самые различные варианты строения конечного мозга: от инвертированного у хрящевых и двоякодышащих, до эвертированного у лучеперых.
Структура полушарий у изученных хрящевых рыб чрезвычайно разнообразна. У многих акул и скатов парность полушарий внешне выражена только в самой ростральной части мозга (см. рис. 5). Выделяются виды, полушария которых имеют хорошо развитые латеральные желудочки (например, у акулы Squilus acanthias), и виды, у которых они практически редуцированы (например, у акулы Mustelus canis или ската Raja clavata).
Цитоархитектоника и состав конечного мозга подробно изучены только у некоторых представителей хрящевых рыб (в основном пластиножаберных). В паллиуме конечного мозга акулы Squalus acanthias выделяют три отдела: медиальный, дорсальный и латеральный, которые гомологичны соответствующим отделам паллиума конечного мозга инвертированного типа других позвоночных (рис. 84). У данного вида акул наиболее дифференцированным отделом паллиума является дорсальный, в пределах которого, впервые у позвоночных, формируется корковая пластинка - слой клеток, отделенных от перивентрикулярной зоны. В ней обнаружены высокодифференцированные пирамидоподобные и звездчатые короткоаксонные нейроны, сходные по структуре с аналогичными клеточными элементами корковой пластинки паллиума высших позвоночных.
В каудальной части конечного мозга дорсальный отдел полушария представлен так называемым центральным ядром. Медиальный
паллиум занимает большую часть дорсомедиальной стенки полушария и разделяется на две зоны: медиальную (M1) и дорсомедиальную (М2). Медиальная зона наиболее выражена в ростральных отделах полушария, дорсомедиальная - в средних и каудальных отделах полушария, отличается ют зоны M1 присутствием крупных нейронов радиального типа.
В латеральном паллиуме нейроны мигрируют в толщу стенки полушария, однако в отличие от дорсального паллиума не формируют корковой пластинки. Эта часть полушария представлена популяциями изодендритных нейронов лофодендритного и радиального типов. У некоторых хрящевых рыб, в частности у акулы Squalus acanthias, в каудальных отделах латерального паллиума, на границе со стриатумом, выделяют группу нейронов - ядро А, которое рассматривается как гомолог паллиальной части амигдалы.
Субпаллиальные отделы конечного мозга хрящевых рыб значительно развиты и представлены стриатумом, обонятельным бугорком, септумом и амигдалой (рис. 84). У многих изученных акул стриатум разделяется на дорсальный и вентральный отделы, которые отличаются сложным нейронным составом и по структуре и гистохимическим особенностям (связи изучены недостаточно) сравниваются с компонентами стрио-паллидарной системы высших позвоночных. Септум занимает медиовентральную часть стенки полушария и разделяется на медиальное и латеральное ядра.
У скатов, среди которых наиболее изучены представители отрада Rajaformes, структура полушарий резко отличается от таковой у многих акул. Так, латеральные желудочки практически редуцированы, а паллиум и субпаллиум представлены диффузными клеточными массами, среди которых выделить какие-либо отделы крайне трудно.
Интересно отметить, что на ранних стадиях развития нервной системы ската Raja clavata в полушариях конечного мозга формируются зачатки паллиальных формаций и есть отчетливо выраженные желудочки. Только к 71-73 суткам развития в паллиуме и субпаллиуме начинается активная миграция клеток и полушария приобретают свойственную взрослым формам диффузную структуру. Учитывая то, что скаты, как полагают, произошли в эволюции от акулоподобных предков, данные преобразования структуры конечного мозга можно рассматривать как вторичные.
Исследование афферентных связей конечного мозга пластиножаберных рыб показало, что зона прямых обонятельных проекций ограничена рядом отделов полушария (переднее обонятельное ядро, латеральный паллиум, обонятельный бугорок, часть стриатума, септум). В корковом слое дорсального паллиума зарегистрированы ответы на световые и обонятельные воздействия, центральное же ядро является местом окончания таламических
афферентов, приносящих в полушарие информацию о зрительных раздражителях. Эти проекции топографически обособлены: вентролатеральное ядро таламуса, связанное с сетчаткой, проецируется в среднюю часть центрального ядра; латеральное ядро, связанное с тектумом, проецируется в каудальные отделы. Это свидетельствует о формировании у некоторых хрящевых рыб двух самостоятельных систем связей: ретино-таламо-теленцефальной и ретино-текто-таламо-теленцефальной. В центральное ядро, а также в обонятельный бугорок поступают афференты от таламических ядер, связанных с мозжечком и органами боковой линии.
Эфференты изучены недостаточно подробно. Они берут начало как в паллиальных, так и в субпаллиальных отделах полушария и адресованы обонятельным луковицам, таламусу, нижней доле гипоталамуса, стволу и верхним сегментам спинного мозга.
Основная масса проекций проходит в составе двух крупных трактов: медиального и латерального пучков переднего мозга. Комиссуральные волокна идут в основном в составе передней и задней комиссур конечного мозга, расположенных в средних иг каудальных отделах полушарий.
В целом, среди современных хрящевых рыб имеется значительная вариабельность в организации одноименных структур конечного мозга, а у некоторых акул уровень его структурно-функциональной организации по ряду параметров (структурных, физиологических, биохимических) сравним с таковым у высших позвоночных.
Структура конечного мозга двоякодышащих и кистеперых рыб изучена далеко неполно. Полушария развиваются путем инверсии, однако их организация у представителей двух основных групп Dipnoi значительно различается. У Neoceratodus forsteri практически отсутствует медиальная стенка полушария, а на ее месте в ростральных отделах мозга развивается мембранозная оболочка, septum ependimale, лишенная нейронов, тогда как у Protopterus dotloi медиальная стенка хорошо выражена, и полушария имеют типичное строение. Несмотря на то, что паллиум двоякодышащих рыб развит относительно слабо и уступает по размеру субпаллиуму, в его дорсальном отделе формируется, как и у некоторых хрящевых рыб, корковая пластинка. Среди субпаллиальных структур выделяют стриатум, септум, обонятельный бугорок, нейронный состав и связи которых практически не изучены.
Инвертированный мозг единственного в современной фауне представителя кистеперых рыб - латимерии - имеет необычное строение: в ростральных отделах конечного мозга дорсолатеральная стенка полушария сильно разрастается и представлена диффузной массой клеток. В средних и каудальных отделах в паллиуме выделяют медиальный, дорсальный и латеральный
участки, клетки которых сосредоточены в основном в перивентрикулярной зоне. Отделы субпаллиума (стриатум, септум, обонятельный бугорок) дифференцированы слабо.
Конечный мозг лучеперых рыб в силу особенностей его формирования в эмбриогенезе значительно отличается от такового других позвоночных (см. рис. 4). В эвертированных полушариях выделяют две основные области: дорсальную (D) и вентральную (V), разделяемые в свою очередь на зоны.* У представителей различных групп лучеперых рыб степень дифференцировки полушарий значительно варьирует, однако четко прослеживается эволюционная тенденция усложнения их структуры у представителей высших отрядов по сравнению с низшими (рис. 85). Так, у многопера Polypterus ornatipinnis дорсальный отдел полушария не дифференцирован на зоны, а основная масса нейронов сосредоточена в узком субэпендимном слое под поверхностью полушария. Вентральная область разделяется на три зоны: дорсальную (Vd), вентральную (Vv) и латеральную (VI). У хондростей и голостей, наиболее изученными представителями которых являются осетровые и ильные рыбы, в дорсальной области уже дифференцируются три или четыре зоны: медиальная (Dm), дорсолатеральная (Dd+l), разделяемая часто на дорсальную (Dd) и латеральную (D1), и центральная (Dc). Нейроны активно мигрируют в толщу стенки полушария, формируя в ряде случаев (осетры) слоистые структуры.
У костистых рыб встречаются различные варианты строения паллиума и субпаллиума: от слабодифференцированных у низших, до сложнейшего комплекса у представителей высших костистых рыб, где в составе полушария выделяют до полутора десятков отдельных зон.
Характер связей конечного мозга лучеперых рыб изучен далеко не полностью. Однако совершенно очевидно, что, с одной стороны, зона окончания обонятельных афферентов охватывает не весь конечный мозг, а с другой - ряд областей и зон полушария, как получающих обонятельные входы, так и лишенных их, имеют проекции других сенсорных систем. Так, у костистой рыбы Sebasticus marmoratus обонятельные афференты проецируются билатерально в каудальный отдел дорсальной области - зону Dp, часть зоны DC и ряд районов вентральной области (зоны VI, Vi, Vs). В зоны Dm и Dd приходят афференты из преоптической области и ядер вентрального таламуса, вероятно связанного с акустико-латеральной системой. Зона D1 получает волокна от ядер таламуса, связанных с тектумом. Из стволовой части мозга (в частности, из ядер шва и голубого пятна) афферентные волокна билатерально достигают зон Dc, Dd и дорсальной части медиальной зоны.
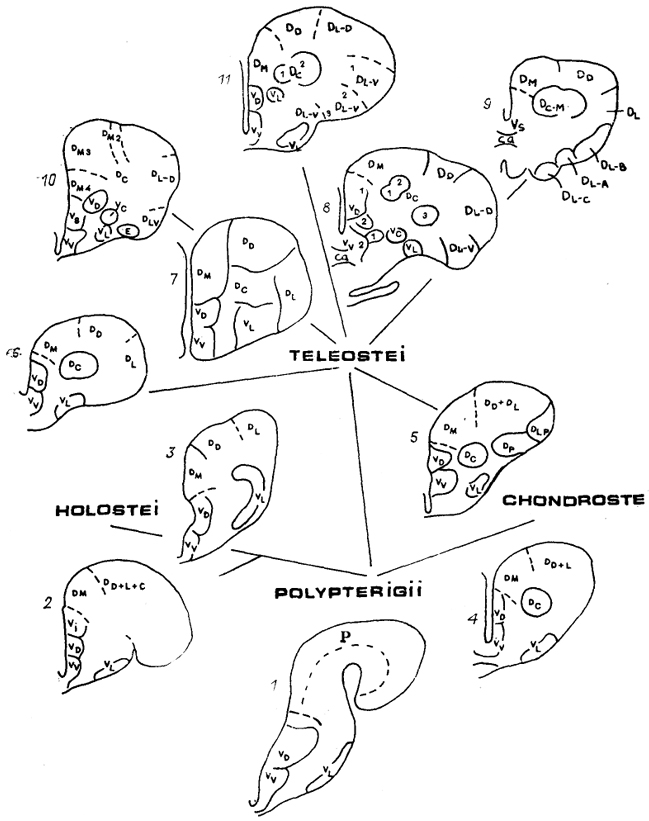
Рис. 85. Схема строения полушарий конечного мозга разных лучеперых рыб: многоперы (1), грлостеи (2, 3), хондростеи (4, 5), низшие (6, 7) и высшие 8-11) костистые.
Буквами обозначены зоны полушарий по классификации R. Nieuwenhuys.
Эфферентные связи полушария, проходящие в основном по латеральному пучку переднего мозга, образованы аксонами нейронов зон DC, Dd и Dm. Они проецируются ипсилатерально в таламус, нижнюю долю гипоталамуса, средний мозг. Через медиальный обонятельный тракт центрифугальные волокна из зон Dp, DC, Vd достигают обонятельной луковицы.
Внутренние связи полушарий обеспечивают взаимодействие между их областями и зонами. Так, зона DC ипсилатерально связана с соседними зонами паллиума, а через переднюю комиссуру с зоной D1 соседнего полушария. Отмечены двусторонние связи медиальной зоны. Дорсальная зона паллиума посылает .многочисленные волокна в медиальную зону, откуда берут начало эфференты к нижней доле гипоталамуса.
Сходные результаты получены при изучении связей конечно-то мозга и других представителей лучеперых рыб.
В связи с особенностями эмбриогенеза конечного мозга лучеперых рыб сложным является вопрос гомологии их полушарий со структурами конечного мозга инвертированного типа. Одни исследователи предлагают проводить прямую гомологию зон :мозга двух типов, другие считают невозможным проведение каких-либо сравнений, поскольку рассматривают конечный мозг .лучеперых рыб как особую линию в эволюции мозга позвоночных. Третьи, а их большинство, основываясь на современных данных о морфологии, гистохимии и связях конечного мозга позвоночных и учитывая особенности процессов инверсии и эверсии, считают, что можно найти решение данной проблемы. Так, зона Dp рассматривается ими как возможный гомолог латерального паллиума, зоны D1 и Dd - медиального и дорсального паллиума соответственно. Каудальные отделы зоны Dm и часть зоны DC сравнивают со стриатумом, отмечая при этом, что другие части этих зон включают компоненты дорсального паллиума. Ростральные отделы медиальной зоны гомологизируются с частью амигдалы. Вентральные зоны Vd и Vv сравниваются с областью септальных ядер, зона VI - с обонятельным бугорком, а каудальные отделы Vs, Vp, Vi- с частью амигдалы (рис. 86).
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|