- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Модульный уровень 20 страница
Особое место в системе субпаллиальных образований мозга птиц занимают парольфакторная область, lobus parolfactorius, и базальное ядро, nucl. basalis (рис. 89). Парольфакторная область расположена в вентромедиальной части полушария и граничит с полостью мозгового желудочка. Ее часто рассматривают как часть РР, однако ее нейроны не проецируются в РР, образуют связи с ядром ТРС. Скорее всего это самостоятельное-образование субпаллиума птиц.
Базальное ядро расположено в вентральном отделе полушария и имеет обширные связи с различными структурами головного мозга. Оно получает проекции от ядер тройничного нерва и латеральной петли, обонятельного бугорка, периферической части эктостриатума, получающего зрительную информацию. Таким образом, базальное ядро является зоной конвергенции различной сенсорной информации, однако необычно то, что сенсорные проекции из ствола мозга достигают этого отдела полушария, минуя таламус. Исследователи рассматривают данную структуру как уникальную для птиц и связывают ее функцию с координацией процесса клевания.
Обонятельный бугорок, септум и nucl. accumbens мозга птиц практически не изучены (за исключением общей цитоархитектоники, сходной с таковой у рептилий).
Архистриатум, archistriatum (поле К), конечного мозга птиц, сравнивается с амигдалярным комплексом мозга других позвоночных. Он отчетливо выражен в мозге многих птиц и достигает наибольшего развития у пингвинов. Архистриатум разделяется на базолатеральную и кортикомедиальную части. Довольно крупные радиальные нейроны архистриатума располагаются одиночно. Ростральные отделы образуют связи с таламусом, тектумом, ретикулярной формацией, ядрами ствола; каудальные - с гипоталамусом и гиппокампом.
Таким образом, субпаллиальные отделы конечного мозга птиц имеют черты организации как специфические, так и общие
для всех амниот. Это касается формирования двусторонних связей стриатума с паллиальными формациями, с одной стороны, и с диенцефало-мезенцефалическими центрами - с другой.
Конечный мозг млекопитающих обладает наиболее сложными корковыми формациями полушарий. Основу цитоархитектонического деления коры полушарий разработал в начале XX в. К. Бродман. Он разделил кору на две основные области: гомогенетическую, для которой характерно шести-семислойное строение, и гетерогенетическую, где такое строение отсутствует. Гомогенетическая кора разделяется на гомотипическую (шестислойное строение сохраняется и во взрослом состоянии) и гетеротипическую (в процессе эмбриогенеза количество слоев меняется). Каждая из этих территорий разделяется на области, а области на поля, различающиеся по строению, связям, функциональной значимости. Так, у приматов выделено 11 областей, включающих 52 поля.
По классификации И.Н. Филимонова основная область коры, обозначенная им как новая кора (неокортекс, изокортекс) соответствует гомогенетической коре по Бродману. Области старой (архикортекс) и древней (палеокортекс) коры гомологичны только части гетерогенетической коры. Остальную часть коры занимает так называемая межуточная кора (мезокортекс), состоящая из периархикортекса, отделяющего новую кору от старой, и перипалеокортекса, отделяющего новую кору от древней.
В состав новой коры, по мнению И.Н. Филимонова, должен быть включен весь паллиум, в пределах которого стенка мозга уже на самых ранних этапах развития характеризуется наличием основных слоев Гисса (матрикс, промежуточный слой, корковая пластинка, краевой слой). Старая и древняя кора, в отличие от новой, на ранних стадиях развития характеризуется неполнотой строения стенки мозга. Межуточная кора имеет переходный тип строения. К палеокортексу относятся пириформная кора (PC) и препириформная область полушария, а также обонятельная луковица, переднее обонятельное ядро, обонятельный бугорок, септум, часть амигдалы и ряд мелких ядер вентромедиальной стенки полушария.*
В состав архикортекса у млекопитающих включают районы гиппокампа (Н), зубчатой фасции (FD), субикулума (S). Пе-риархикортекс представлен пре- и парасубикулумом (PrS, PaS), taenia tecta (TT), энторинальной (ERC) и цингулярной (СС) корой; перипалеокортекс - переходной инсулярной областью (CI). Исследование большого числа млекопитающих показало, что деление коры на отмеченные выше основные области
всюду сохраняет свое значение и проблема их гомологии у представителей разных групп млекопитающих не встречает существенных затруднений.
В эволюции млекопитающих новая кора благодаря своему интенсивному росту оттеснила старую и древнюю кору на медиальную и вентральную поверхности полушария. В результате сформировались две основные борозды: гиппокамповая (sh), отделяющая старую кору от новой, и ринальная (srh), отделяющая древнюю кору от новой. Эти борозды отчетливо выражены в конечном мозге всех млекопитающих. Далее происходило образование борозд и извилнн в пределах корковых областей. По степени развития борозд и извилин полушария мозга млекопитающих разделяются на два типа: лиссэнцефаличеекий, для которого характерно отсутствие или очень слабое развитие борозд на поверхности полушария, и гирэнцефалический - с более или менее развитой системой борозд и извилин. Первый тип характерен для однопроходных, низших сумчатых, насекомоядных, рукокрылых, грызунов. Остальные млекопитающие имеют на поверхности полушария систему борозд и извилин. Вместе с тем необходимо подчеркнуть, что среди разных групп современных млекопитающих есть представители, анатомия мозга которых отличается от характерной для всей группы. Так, у ехидны полушария имеют необычно развитую систему борозд и извилин, захватывающих даже область древней коры (рис. 91).
Межвидовая и межгрупповая изменчивость проявляется в варьировании многих важных показателей уровня развития и дифференцировки коры: площади и объема одноименных областей и полей, толщины коры, степени выраженности и толщины отдельных слоев корковой пластинки, размеров и плотности расположения нейронов.
В процессе эволюции млекопитающих происходило последовательное увеличение площади, занимаемой ассоциативными полями и усиление вариабельности в расположении одноименных зон коры у разных животных (рис. 92).
Каждое поле и каждая область коры имеют многочисленные связи с другими отделами коры и частями мозга. Примерно половина из них образована афферентными волокнами. Они подразделяются на две большие группы: кортико-кортикальные (ассоциативные и межполушарные) и образованные волокнами из нижележащих отделов мозга.
Ассоциативные связи объединяют поля в пределах одного полушария. В образовании этих связей принимают участие нейроны многих слоев коры. Количество, типы и характер распределения по слоям нейронов, образующих ассоциативные проекции, зависят от поля коры, вида животного и, как полагают, могут меняться в процессе онтогенеза. Пирамидные нейроны образуют от 60 до 90% таких связей. Число и распределение
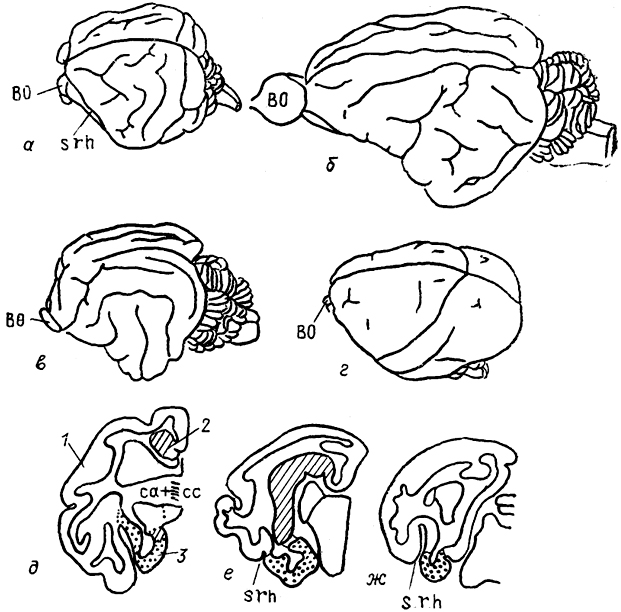
Рис. 91. Внешний вид головного мозга различных млекопитающих (а-г) в поперечные срезы мозга ехидны (д-ж) (Welker, Lende, 1980).
а - ехидна, б - грызун капибара, в - кошка, г - беличья обезьяна; 1-3 - отделы коры: неокортекс (1), архикортекс (2), палеокортекс (3).
терминалей ассоциативных волокон зависят от слоя коры, в котором проходят кортико-кортикальные аксоны, а также от типа полей, связанных между собой. Корковые поля образуют в пределах полушария связи с разным количеством полей.
Комиссуральные волокна сосредоточены в основном в мозолистом теле, corpus callosum. Его формирование связано с развитием новой коры, однако при резком увеличении у высших млекопитающих, по сравнению с низшими, площади мозолистого тела, плотность волокон сохраняется на одном уровне: примерно 400 тысяч волокон на 1 мм2 сечения. В коре есть участки, как образующие комиссуральные связи (их большинство), так и лишенные их (поле 17 зрительной коры). Ней-
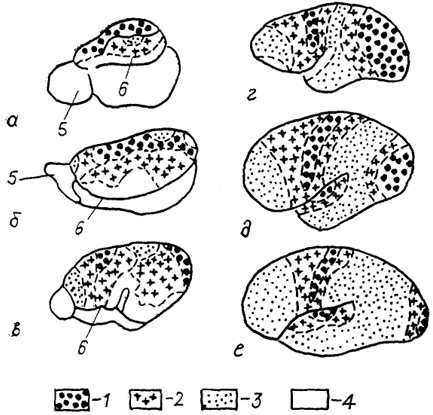
Рис. 92. Расположение и степень развития корковых полей различных типов на латеральной поверхности полушарий млекопитающих (Богословская, Поляков, 1981).
а - еж, б - крыса, в - собака, г - мартышка, д - оранг, е - человек; 1-3 - поля леокортекса: первичные (1), вторичные (2) и третичные (3), 4 - территория древней, старой и межуточной коры, 5 - обонятельная луковица, 6 - ринальная борозда.
роны, образующие каллозальные проекции, как и их окончания, располагаются во всех слоях коры.
Афферентные связи включают кроме перечисленных выше таламо-кортикальные и экстраталамические проекции. У разных животных и в разных полях коры характер окончаний таламических афферентов (даже от одноименных ядер таламуса) может варьировать).
Экстраталамические афференты берут начало в основном от центров ствола и в соответствии с источником проекции разделяются на холин-, адрендофамин-, и серотонинергические. Эти волокна проходят во все слои коры (в каждой системе имеются свои особенности распределения окончаний по слоям), идут параллельно поверхности коры на большом расстоянии (до нескольких миллиметров) и образуют как типичные синаптические соединения, так и модуляторные контакты. Общее число таких окончаний составляет от 5 до 12% от числа межнейронных контактов в коре.
Наиболее крупными эфферентными путями считаются кортико-спинальные (пирамидный тракт) и кортико-бульбарные тракты. Они образованы аксонами пирамидных нейронов нижних (V-VI) слоев коры. Большинство эфферентов (особенно идущих от моторных и сенсорных полей коры) имеют строго топическую
организацию. Волокна нисходящих трактов формируют многочисленные коллатерали, оканчивающиеся в различных отделах мозга. Так, коллатерали волокон пирамидного тракта оканчиваются на нейронах красного ядра, образующих руброспинальные проекции, и влияют на их деятельность.
У млекопитающих встречается несколько вариантов строения неокортекса. Первый отмечен у насекомоядных, рукокрылых, грызунов, зайцеобразных. Неокортекс занимает 30-60% общей поверхности коры. Преимущественно развиты нижние слои коры и слой 1. Присутствует большое количество слабодифференцированных, атипичных нейронов. По цитоархитектонике и нейронному составу зоны и поля коры различаются между собой слабо. Среди представителей каждой из этих групп млекопитающих встречаются и крайне специализированные виды, план строения коры которых значительно отличается от такового у всей группы. Так, у ежей обнаружены участки неокортекса, характеризующиеся преимущественным развитием верхних и нижних слоев коры и появлением высокодифференцированных пирамидных и звездчатых нейронов.
Второй тип строения неокортекса встречается у китообразных, конечный мозг которых значительно отличается от мозга других млекопитающих. Он имеет чрезвычайно сложную систему борозд и извилин, неокортекс занимает 97-98% поверхности коры и очень тонкий (0,158-0,202 см), вследствие чего относительный объем неокортекса китообразных оказался меньше, чем у насекомоядных. Цитоархитектоническая и нейронная структура неокортекса имеют многие черты, характерные для мозга низших млекопитающих. При этом в мозге дельфинов обнаружены длинные ассоциативные внутрикорковые связи, развитие которых является специфической чертой конечного мозга приматов. Все это свидетельствует о своеобразном пути эволюции конечного мозга китообразных.
Третий вариант строения неокортекса отмечен у представителей таких отрядов млекопитающих, как хищные, ластоногие, парно- и непарнокопытные, хоботные. В целом их конечный мозг отличается развитием системы крупных дуговых борозд и извилин на латеральной поверхности полушария, сильно затрудняющих сравнение с полушариями конечного мозга других млекопитающих. Неокортекс занимает от 80 до 95% площади коры и имеет большую толщину. Нейронная структура характеризуется увеличением разнообразия и полиморфизма клеток коры, намечается разделение неокортекса на гранулярные и агранулярные поля.
Четвертый тип коры встречается у приматов. В этой группе млекопитающих, филогенетически самой молодой, наиболее высока эволюционная и адаптивная изменчивость. Это проявляется в значительной вариабельности структуры коры в разных полях одного полушария и одноименных полей у разных
животных. Резко увеличивается количество и разнообразие клеточных форм в составе неокортекса, завершается выделение гранулярных и агранулярных областей коры, разрастаются височные доли полушария, большую площадь занимают третичные, ассоциативные поля.
В настоящее время созданы различные теории организации неокортекса млекопитающих, среди которых наибольшее распространение получила модульная концепция структурно-функциональной организации коры. Интересную модель эволюции модулей в коре мозга млекопитающих предложил П. Морган.
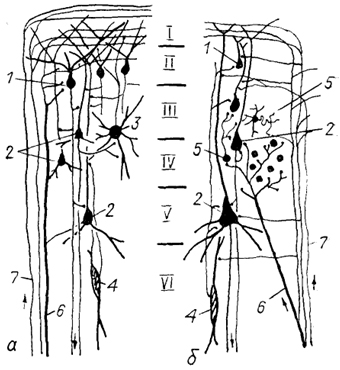
Рис. 93. Схема возможной эволюции модульного строения коры млекопитающих (Glezer e. а., 1988).
а - исходный гипотетический модуль, б - модифицированный модуль. 1 - пирамиды II слоя с широкой зоной ветвления дендритов, 2 - пирамиды III-V слоев, 3 - мультиполярная клетка, 4 - биполярная клетка, 5 - короткоаксонные звездчатые нейроны разных типов, 6 - специфическиеи 7 - неспецифические таламо-кортикальные афференты.
Согласно этой модели в эволюции происходило образование в коре модулей с их последующей модификацией (усложнением) и увеличением количества. Исходный (гипотетический) тип мозга предков млекопитающих характеризовался наличием в: коре небольшого количества просто организованных модулей. В этих модулях специфические и неспецифические таламо-кортикальные волокна, а также ассоциативные и комиссуральные проекции направлялись преимущественно в I слой, контактируя с апикальными дендритам и пирамидных и непирамидных нейронов (рис. 93). Большие мультиполярные изодендритные нейроны посылали свои аксоны во второй слой, обеспечивая внутрикорковые связи. Таким образом, основным "афферентным" слоем коры был II слой с его специализированными пирамидными клетками, аксоны которых направлялись во все нижележащие слои коры.
В процессе эволюции в модулях происходило разделение зон окончаний основных афферентных проекций. Таламо-кортикальные афференты оканчивается преимущественно в новом, интенсивно развивающемся IV слое, где появляется большое число высокоспециализированных звездчатых нейронов. Другие афференты направляются в верхние слои коры, давая на своем протяжении многочисленные коллатерали.
В разных группах млекопитающих процесс модификации и увеличения количества модулей шел, по мнению автора этой гипотезы, независимо, в результате чего к настоящему времени сформировалось четыре основных типа неокортекса. Консервативный, при котором сохраняются многие признаки гипотетического типа коры, встречается у представителей современных "отрядов насекомоядных и рукокрылых. Прогрессивно-консервативный тип, характеризующийся некоторым увеличением количества модулей и незначительным их усложнением. Животные с таким типом неокортекса встречаются практически во всех отрядах современных млекопитающих (например, лемуры и мартышки среди приматов). При прогрессивном типе строения •коры количество, степень сложности и разнообразие модулей значительно увеличивается. В каждом отряде млекопитающих есть представители, неокортекс которых имеет черты прогрессивного типа (среди приматов это высшие обезьяны и человек). Консервативно-прогрессивный тип отличается резким увеличением количества модулей в коре при незначительной их модификации, что приводит к формированию "монотонной" структуры коры (китообразные).
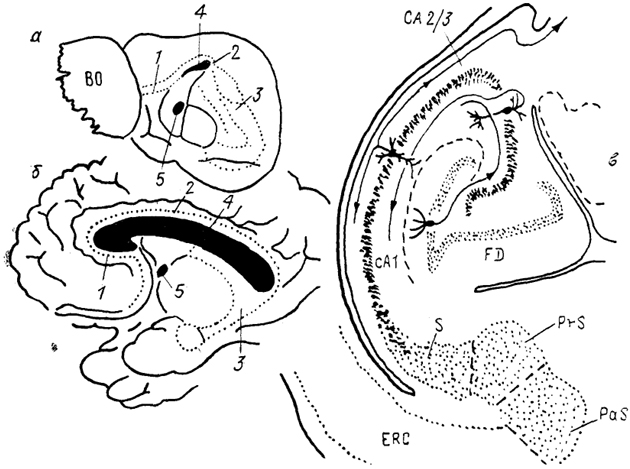
Рис. 94. Организация гиппокампа у ежа (а) и человека (б) и строение гиппокампа лемура на тангенциальном срезе (в) (Stephan, Malonescu, 1980).
1 - прекаллозальный, 2 - супракаллозальный и 3 - посткаллозальный отделы гиппокампа, 4 - мозолистое тело, 5 - передняя комиссура, СА 1 и СА 2/3 - отделы гиппокампа, ERC - энторинальная кора, FD - зубчатая фасция, PaS - парасубикулум, PrS - пресубикилум, S - субикулум.
Старая кора (архикортекс) мозга млекопитающих образована в основном структурами гиппокампа, который разделяется
у плацентарных млекопитающих на три отдела: прекомиссуральный (прекаллозальный), супракомиссуральный (супра-каллозальный) и ретрокомиссуральный (посткаллозальный). Из-за сильного развития у высших млекопитающих мозолистого тела первые два отдела у них редуцируются (рис. 94). Прекомиссуральный отдел, расположенный между зоной переднего обонятельного ядра и мозолистым телом, представлен мелкими пирамидными клеткамл и связан афферентными и эфферентными проекциями с пириформной и энторинальной корой. Средняя часть гиппокампа превращается в узкую полоску серого вещества, связывающего передний и задний отделы гиппокампа. Ретрокомиссуральный отдел наиболее развит и представлен структурами субикулума (S) и собственно гиппокампа, который состоит из ряда зон (СА 1-4) и зубчатой фасции (FD). Гиппокамп имеет корковую структуру и состоит из пяти слоев, основным из которых является III слой, образованный пирамидными нейронами. У низших млекопитающих этот слой узкий, компактный; у высших - широкий, диффузный.
Гиппокамп - важнейший компонент лимбической системы конечного мозга, связанный со многими отделами головного мозга. Он получает афферентные проекции от различных зон неокортекса (включая ассоциативные поля), а также септума, амигдалы, ряда ядер таламуса, многих отделов ствола мозга. Эфферентные проекции гиппокампа образованы аксонами исключительно пирамидных нейронов и проходят в основном в составе свода. Через него обеспечивается связь гиппокампа с амигдалой, таламусом, гипоталамусом, палеокортексом, септумом. У высших млекопитающих над мозолистым телом формируется пучок волокон - пояс, cingulum, через который гиппокамп связан с другими отделами архикортекса и рядом полей неокортекса. Комиссуральные связи осуществляются через комиссуру гиппокампа.
Древняя кора (палеокортекс) у млекопитающих занимает в полушарии вентральное положение, оттесненная с латеральной поверхности сильно разросшейся новой корой. Для нее характерны слабая слоистость и неполное отделение от соседних клеточных масс стриатума. Основная часть палеокортекса - пириформная кора - получает обонятельные проекции по латеральному обонятельному тракту, волокна которого оканчиваются на апикальных дендритах пирамидных нейронов пириформной коры. Аксоны этих нейронов направляются в обонятельный бугорок, а также в префронтальную область коры. Палеокортекс имеет эфферентные связи с амигдалой и эпиталамусом. Обращает на себя внимание тот факт, что у животных, имеющих редуцированную обонятельную систему (например, дельфины, некоторые птицы), палеокортекс развит хорошо. Это свидетельствует о том, что палеокортекс функционально связан не только с обонянием.
Субпаллиальные формации, конечного мозга млекопитающих включают в себя комплекс ядер: хвостатое ядро, nucl. caudatus, и скорлупу, putamen, часто рассматриваемые как единая структура (неостриатум), а также бледный шар, globus pallidus, палеостриатум, ограду, claustrum, амигдалу, структуры вёнтромедиальной стенки полушария (септум, обонятельный бугорок, ядро диагональной связки Брока, безымянная субстанция, nucl. accumbens).
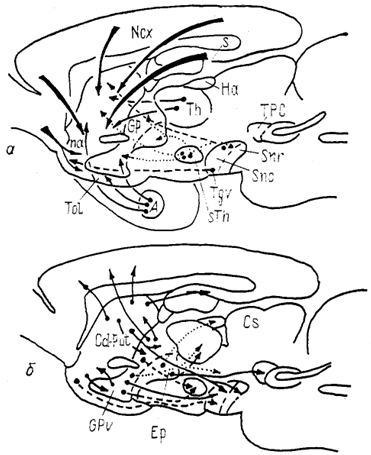
Рис. 95. Основные связи стрио-паллидарной системы млекопитающих на примере мозга крысы (Heimer, Alheid, Zaborzsky, 1985).
а - афферентные, б - внутренние и эфферентные связи.
Цитоархитектоническая и нейронная организация стриатума млекопитающих (он включает формации нео-, палеостриатума и ограду) очень сложна. Неостриатум цитоархитектонически неоднороден. Его клетки образуют сложные пространственные комплексы-стриосомы, окруженные более рыхло расположенными нейронами, составляющими так называемый матрикс. Выделено несколько типов нейронов, различающихся по размеру (крупные, средние, мелкие), по типу аксонов (длинно- и короткоаксонные) и характеру дендритных ветвлений (густо- и редковетвистые; шипиковые и бесшипиковые). Показано, что клетки любого размера могут быть как длинноксонными, образующими эфферентные проекции, так и короткоаксонными, фомирующие внутристриарные связи. Крупные длинноаксонные нейроны дают восходящие проекции в кору, тогда как мелкие и средние длинноаксонные нейроны - нисходящие проекции. В эволюции млекопитающих доля мелких нейронов разного типа увеличивается.
Бледный шар разделяется цитоархитектонически на внутренний и наружный сегменты, нейронный состав которых довольно однообразен. Это в основном крупные редковетвистые нейроны ретикулярного типа со строго упорядоченной ориентацией дендритов,
образующих систему микромодулей в виде дисков, параллельных друг другу и латеральной границе ядра, но расположенных перпендикулярно направлению хода аксонов нейронов неостриатума. Таким образом, аксоны клеток хвостатого ядра и скорлупы, проходя в бледный шар, пронизывают сотни дендритных модулей и образуют сложные синаптические комплексы.
Стриатум млекопитающих имеет сложную систему связей с большим числом мозговых структур (рис. 95). Основной тенденцией в организации афферентных и эфферентных проекций стриатума является формирование кольцевых связей: неокортекс → неостриатум → неокортекс, неокортекс → неостриатум → палеостриатум → таламус → неокортекс, неостриатум → палеостриатум → неостриатум, неостриатум → палеостриатум → таламус → неостриатум, стриатум → черная субстанция → стриатум. Многие из этих связей топографически упорядочены. Например, в случае корково-неостриатарных связей нейроны глубоких слоев коры (V, VI) из определенной зоны неокортекса проецируются в стриосомы, верхних (II, III) - в матрикс. При этом входы из различных полей коры в неостриатуме могут перекрываться, что создает основу для конвергенции на нейронах стриатума разнообразной информации.
Таким образом, филогенетически молодая система связей стриатума с корковыми формациями получает у млекопитающих дальнейшее развитие, обеспечивая ему роль одного из интегративных центров конечного мозга.
Ограда - тонкая полоска серого вещества, отделенная от скорлупы пучками волокон, образующих наружную капсулу, capsula externa, а от коры ее отделяет узкая полоска белого вещества - самая наружная капсула, capsula extrema. Для нейронного строения ограды характерна значительная полиморфность. Наиболее многочисленны корковые выходы из различных зон неокортекса. Афференты также поступают из стриатума, амигдалы, таламуса и черной субстанции. Эфференты ограды направляются к новой коре, стриатуму, амигдале, обонятельной луковице, таламусу и мозжечку. Есть данные, что ограда имеет корковое происхождение.
В септальной зоне конечного мозга млекопитающих, включающей медиальное и латеральное септальные ядра, ядро диагональной связки Брока, nucl. accumbens, и функционально связанной с лимбической системой, появляется, в отличие от аналогичной зоны конечного мозга рептилий и птиц, большое количество высокоспециализированных короткоаксонных нейронов. Однако, анализ связей, структуры и гистохимических характеристик позволяют проводить гомологию основных отделов септальной зоны мозга рептилий, птиц и млекопитающих.
Обонятельный бугорок имеет слоистое строение (различают наружный слой волокон, средний слой пирамидоподобных нейронов
и внутренний слой полиморфных клеток). Степень выраженности слоев у разных видов млекопитающих значительно варьирует. Афферентные проекции поступают из обонятельной луковицы, пириформной коры, амигдалы, преоптической области, ряда зон неокортекса, интраламинарных ядер таламуса, черной субстанции. Эфференты направляются в септальную зону и далее в гиппокамп, стриатум, бледный шар, дорсомедиальную область таламуса и эпиталамус, амигдалу и пириформную кору. Таким образом видно, что обонятельный бугорок млекопитающих связан не только с обонятельной, но и с лимбической и стриопаллидарной системами.
Амигдала млекопитающих состоит из большого числа (до 10) ядер, которые объединяются в несколько групп: базолатеральную (латеральное, основное и добавочное ядра) - наиболее филогенетически молодую часть амигдалы, связанную с корковыми формациями; кортикомедиальную (периамигдаляр-ная кора, среднее и заднее кортикальные ядра, медиальное ядро) - филогенетически более старую часть, связанную с обонятельной системой; центральную (центральное ядро, переднеамигдалярная зона) - связанную с гипоталамусом и ядрами ствола мозга, контролирующими вегетативные функции организма. В филогенезе млекопитающих в амигдале происходило два процесса. Во-первых, уменьшалась доля, занимаемая ядрами кортикомедиальной группы, и увеличивался объем базолатеральной группы ядер (последняя достигает наибольшего развития и максимальной сложности у приматов и человека). Во-вторых, у высших млекопитающих вследствие сильного развития новой коры амгидала смещается с вентральной стенки полушария на вентромедиальную. Многие вопросы организации амигдалярного комплекса конечного мозга высших позвоночных еще не решены.
Основной вопрос, который возникает при изучении конечного мозга, касается гомологии его структурных подразделений у представителей разных групп позвоночных.
В паллиальных отделах конечного мозга медиальный паллиум и часть дорсального паллиума мозга рептилий рассматриваются как гомологи медиодорсальной кортикоидной пластинки птиц и структур архикортекса млекопитающих; ДВК и часть дорсальной коры рептилий - гипер- и неостриатуму птиц и неокортексу млекопитающих; латеральный паллиум рептилий сравнивается с дорсолатеральным паллиумом птиц и пириформной корой млекопитающих. В субпаллиуме возможными гомологами считаются такие структуры как: неостриатум рептилий - палеостриатум возрастающий птиц - стриопаллидарный комплекс (хвостатое ядро и скорлупа) млекопитающих, а также палеостриатум рептилий - часть палеостриатума примитивного птиц и бледный шар млекопитающих.
Проведение сравнений между более мелкими подразделениями
крайне затруднительно, поскольку развитие паллиума и субпаллиума (и их отдельных образований) шло в разных линиях высших позвоночных независимо и параллельно, благодаря чему гомологичные отделы могли приобрести разную структурную организацию.
* В настоящее время исследователи на основании гистохимических, ходологических и эмбриологических данных относят перечисленные здесь структуры (начиная с обонятельного ядра) к субпаллиальным формациям.
ГЛАВА 9
ФУНКЦИОНАЛЬНЫЕ АСПЕКТЫ ПРЕОБРАЗОВАНИЯ НЕРВНОЙ СИСТЕМЫ
В ряду проблем, которые решает эволюционная морфология, вопросы преобразования нервной системы занимают значительное место и представляются наиболее сложными. Это обусловлено целым рядом объективных причин: значительным разнообразием модификаций мозга даже у представителей близких таксонов, невозможностью получения данных о строении нервной системы вымерших форм, а также неразработанностью критериев, по которым можно было бы оценивать уровень ее эволюционного развития. Как и любую из морфофункциональных систем, нервную систему можно анализировать с позиций эволюционной гистологии, анатомии, биохимии.
Так, традиционно полагают, что наиболее высокий уровень организации нервной ткани - ламинарный, перивентрикулярное расположение клеточных масс рассматривается как более примитивное, сложились представления и о разной степени дифференциации нервных элементов. Таким образом, эволюционная гистология оперирует определенными критериями оценки уровня организации нервной ткани (нейронного, синаптического, в последнее время - модульного).
Эволюционно значимым признаком является возрастание массы мозга по отношению к массе тела. Существуют и биохимические признаки, по которым можно судить об уровне организации мозга.
Эволюционная физиология часто использует все эти признаки в комплексе с оценкой сложности поведения.
Несмотря на значительный объем данных, полученных в этих исследованиях, судить о преобразовании мозга в рамках общей теории эволюции еще очень трудно.*
Непосредственное перенесение некоторых представлений о морфогенезах на развитие мозга привело к формированию точки
зрения, согласно которой в процессе эволюции происходит постепенное добавление его отделов, которые благодаря новым связям с ранее образовавшимися отделами, "подчиняют" их себе. Этот же подход обнаруживается и в попытках оценить преобразование мозга в эволюции с позиции преемственности основных функциональных систем ЦНС.
Ставшая наиболее известной классификация такого рода была выдвинута в 1908 г. Л. Эдингером, который рассматривал мозг позвоночных как совокупность двух фундаментальных единиц - "нового", neencephalon, и "древнего" мозга, paleencephalon. Последний является составляющей мозга у всех позвоночных, включая человека и единственной у круглоротых и рыб. Его деятельность обеспечивает локомоцию, репродуктивное поведение и другие инстинкты. Новый мозг появляется лишь у наземных позвоночных и, постепенно увеличиваясь, достигает наибольшего развития у млекопитающих, Он обеспечивает сложные формы их поведения, в том числе обучение и память. У других высших позвоночных его развитие приводит к расширению "поведенческого репертуара".
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|