- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Вегетативная нервная система 4 страница
Л. А. Орбели выявил адаптационно-трофическую роль мозжечка.Нейроны зубчатой фасции мозжечка оценивают состояние внутренних органов и приспосабливают их деятельность, так же как и деятельность мышц, к интересам целого организма.
Мозжечок контролирует артериальное давление и дыхание при движениях, а также оказывает влияние на тонус гладкой мускулатуры кишечника. При повреждении мозжечка нарушается углеводный, минеральный и белковый обмен, а также процессы энергообразования, терморегуляции и кроветворения. Мозжечок участвует в репродуктивных функциях.
Благодаря обширным связям мозжечка с ретикулярной формацией среднего мозга, красными и вестибулярными ядрами, гипоталамусом, лимбическими структурами и корой больших полушарий он выступает в роли своеобразного компаратора, динамически оценивающего движения и деятельность внутренних органов организма (см. также разд. 7.6).
3.11. ФИЗИОЛОГИЯ ГИПОТАЛАМУСА
В гипоталамусе сосредоточены ведущие центры биологических мотиваций голода, жажды, полового влечения, температурные центры, центр сна и бодрствования и др., которые будут подробно рассмотрены в соответствующих разделах учебника. Здесь же расположены многочисленные группы нейронов, продуцирующих факторы, освобождающие гормоны — либерины и статины (см. разд. 4.2 и 6.2).
Мотивациогенные центры гипоталамуса оказывают на кору больших полушарий при мотивациях различного биологического качества специфические восходящие активирующие влияния.
Раздражение ядер переднего отдела гипоталамуса вызывает тро- фотропные эффекты парасимпатической нервной системы. Раздражение задней группы ядер гипоталамуса оказывает эрготропное влияние через симпатическую нервную систему.
3.12. ФИЗИОЛОГИЯ ЛИМБИЧЕСКИХ
ОБРАЗОВАНИЙ ГОЛОВНОГО МОЗГА
Лимбические образования включают: древнюю кору (палеокортекс), старую кору (архикортекс) и срединную кору (мезокортекс).
Палеокортекс объединяет препириформную, периамигдалоид- ную, диагональную кору, обонятельные луковицы, обонятельный бугорок, прозрачную перегородку. Архикортекс включает гиппокамп, зубчатую фасцию, поясную извилину. Мезокортекс объединяет инсулярную кору и парагиппокампальную извилину.
В состав лимбических образований мозга включают миндалины мозга, ядра перегородки, переднее таламическое ядро, мамиллярные тела и гипоталамус (рис. 3.31).
Лимбические структуры получают афферентные возбуждения со стороны практически всех органов чувств:
• обонятельного тракта (перегородка, миндалины);
• зрительных путей через таламус (гиппокамп, инсула);
• слуховых путей (гиппокамп, инсула);
• тактильных путей (инсула);
• от висцеральных органов (инсула).
Лимбические круги.Лимбические образования связаны друг с другом кольцевыми взаимосвязями. Различают несколько лимбических кругов.
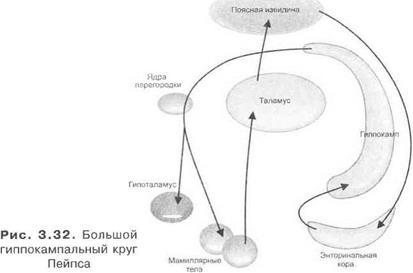
Большой гиппокампальный круг Пейпса. Он включает мамиллярные тела, таламус, гиппокамп, энторинальную кору и снова ядра перегородки, мамиллярные тела гипоталамуса (рис. 3.32).
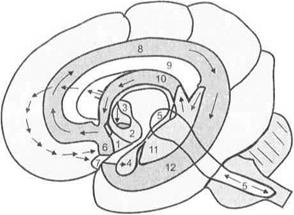
Рис. 3.31.Лимбические структуры головного мозга (по Ф. П. Кос- молинскому):
1 — гипоталамус; 2— мамиллярное тело; 3— передние ядра таламуса; 4 — миндалевидный комплекс ядер; 5— ретикулярная формация ствола мозга; 6— перегородка; 7— гипофиз; 8— поясная извилина; 9— мозолистое тело; 10 — свод; 11 — гиппокамп; 12 — гиппокампова извилина. Светлым обозначена новая кора больших полушарий; темным — лимбические структуры; стрелки указывают направление связей между структурами
Септально-амигдалоидный круг: перегородка — диагональный пучок — миндалина — концевая полоска — перегородка.
Септально-гиппокампальный круг: перегородка — супракало- зальные полоски — гиппокамп — форникс — перегородка.
Перегородка через медиальный пучок переднего мозга связана с варолиевым мостом.
Функции лимбических структур:
• Определяют эмоциональную окраску мотиваций, поведенческих и висцеральных реакций.
• Циркуляция возбуждений по лимбическим кругам определяет суточные ритмы многих физиологических функций: голод, насыщение, мочевыделение, секрецию гормонов корой надпочечников и т.д.
• Участвуют в механизмах памяти. При нарушениях, например, структур гиппокампа с трудом образуются новые навыки, утрачиваются старые навыки, проявляется дефицит кратковременной памяти. Пациенты не узнают лечащих врачей, не помнят дату своего рождения, не могут выучить название больницы и даже не запоминают дорогу к своей кровати. Не помнят счет спортивной игры, теряют нить разговора.
• Участвуют в формировании эпилептической активности, особенно при слабых электромагнитных воздействиях. При этом блокируются болевые ощущения.
3.13. ФИЗИОЛОГИЯ ТАЛАМУСА
Основная функция таламуса — проводниковая. Таламус — коллектор практически всех афферентных путей (за исключением обонятельных), идущих к коре больших полушарий.
В таламусе имеется четкая локализация представительства различных афферентных путей, идущих от различных рецепторов.
Специфические (лемнисковые) проводящие пути к коре больших полушарий распространяются через латеральные ядра таламуса. Эти афферентные пути заканчиваются в определенных специфических ядрах таламуса (передние вентральные, медиальные, вентролатеральные, постмедиальное, постлатеральное, а также медиальные и латеральные коленчатые тела), откуда возбуждения направляются по двум путям: к базальным ядрам конечного мозга и к спеиифиче- ским проекционным зонам коры мозга (зрительной, слуховой, соматосенсорной и др.). К специфическим ядрам таламуса поступает афферентация от тактильных, болевых, температурных, мышечных рецепторов, а также от блуждающих и чревных нервов. Особенно тесно таламус связан с соматосенсорной областью коры большого мозга. Эта взаимосвязь осуществляется благодаря наличию корковоталамических волокон, образующих замкнутые циклические связи. Соматосенсорную область коры и таламус рассматривают как функциональное единое целое. Медиальные и латеральные коленчатые тела — подкорковые центры слуха и зрения соответственно.
Неспецифические ядра таламуса (срединный центр, парацентральное ядро, центральное, медиальное и латеральное, субмедиальное, переднее вентральное, парафасцикулярное, ретикулярное, перивентрикулярное ядро и центральная серая масса) являются местом переключения восходящих активирующих влияний, которые направляются от ретикулярной формации ствола в кору большого мозга.
Специфические и неспецифические ядра таламуса находятся в тесной функциональной взаимосвязи, что обеспечивает первичный анализ и интеграцию всех афферентных возбуждений, поступающих в головной мозг.
Ассоциативные ядра (медиодорсальное, латеральное дорсальное ядро и подушка) обрабатывают информацию различной модальности, которая поступает в 1-е и 2-е слои ассоциативной зоны коры и, частично, в проекционные зоны коры (4-е и 5-е слои).
Высокочастотное раздражение неспецифических медиальных ядер таламуса вызывает локальную кратковременную активацию коры мозга. Низкочастотное раздражение вызывает реакцию «вовлечения» — деактивацию коры (рис. 3.33).
В ядрах таламуса выявлена пространственная топография различных частей тела, в частности сетчатки, языка и кортиева органа.
Функции таламуса:
• определяет двигательные и вегетативные реакции, связанные с сосанием, жеванием, глотанием и смехом;
• участвует в механизмах медленноволновой фазы сна;
• участвует в механизмах боли; определяет возникновение как мучительной каузалгии, так и отсутствие болевых ощущений.
ЭЭГ соматосенсорной коры


Рис. 3.33.Реакция «вовлечения» ЭЭГ соматосенсорной коры при электрическом раздражении срединного центра таламуса
3.14. ФИЗИОЛОГИЯ СТРИАРНЫХ ОБРАЗОВАНИЙ ГОЛОВНОГО МОЗГА
Полосатые тела (corpora striati) состоят из хвостатого ядра и скорлупы (вместе они составляют неостриатум, филогенетически более новое образование), бледного шара (палеостриатум — наиболее древняя часть стриатума) и ограды (рис. 3.34).
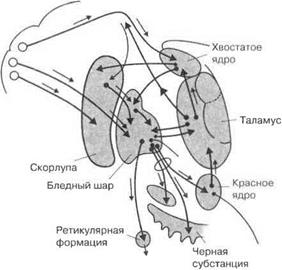
Рис. 3.34.Стриатные образования головного мозга человека и их эфферентные связи
Базальные ганглии связывают кору больших полушарий через черное вещество с таламусом и определяют переход от заданных к выбранным программам поведения. При этом информация снова возвращается по скелетомоторной петле в кору больших полушарий, контролируя силу, амплитуду и направление движения, включая движения мышц и рта. По глазодвигательной петле осуществляется регуляция движения глаз. Возбуждающим медиатором, осуществляющим корково-стриатные связи, является глутамат, а ГАМК между базальными ганглиями и таламусом осуществляет тормозное действие.
Полосатое тело и бледный шар, влияя на нейроны спинного мозга через структуры среднего и продолговатого мозга, координируют тонус и фазовую двигательную активность мышц.
При участии стриатных образований мозга осуществляется синергизм всех элементов таких сложных двигательных актов, как ходьба, бег, лазанье, перешагивание через препятствие, вдевание нитки в иголку; достигаются плавность движений и установка исходной позы для их осуществления.
Полосатое тело не имеет прямых выходов к мотонейронам спинного мозга, а опосредует свои влияния на них через ретикулоспинальный тракт. Эти влияния адресуются к у-мотонейронам спинного мозга, которые регулируют поток проприоцептивных афферентных импульсов, поступающих в спинной мозг от мышечных веретен. Эти афферентные импульсы влияют на возбудимость а- мотонейронов, активность которых определяет рабочее состояние скелетных мышц.
Полосатое тело дает также начало многим двигательным путям экстрапирамидной системы, среди которых выделяют эффекторный путь к паллидуму, далее: красное ядро — руброспинальный тракт — спинной мозг. Бледный шар оказывает тормозящее воздействие на ядра среднего мозга.
Неостриатум вместе с бледным шаром полосатого тела тормозят моторный компонент условных и безусловных реакций организма. Нейроны головки хвостатого ядра принимают участие в организации предпусковых процессов, которые включают перестройку позы, предшествующей произвольным двигательным актам.
Хвостатое ядро, скорлупа и бледный шар участвуют не только в регуляции моторной деятельности, но и в анализе афферентных потоков, в регуляции ряда вегетативных функций, осуществлении сложных форм врожденного пищевого и оборонительного поведения, механизмах кратковременной памяти, а также в регуляции цикла «бодрствование-сон».
При раздражении скорлупы наблюдается слюноотделение и изменение дыхания. При поражении скорлупы наблюдаются трофические поражения кожи.
3.15. ФУНКЦИИ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ
Кортикализация функций.Чем выше животные в эволюционном ряду, тем все большую роль в его функциях берут на себя нейроны коры больших полушарий.
Человек с пораженной корой больших полушарий — глубокий инвалид с утратой речи и целенаправленной деятельности, потерей тонких движений и т.д.
Главная функция коры головного мозга — оценка внутреннего состояния организма и воздействий внешней среды (через аппарат эмоций), а также построение адекватных внутренним потребностям и воздействиям внешней среды форм поведения и их оценка.
Кора больших полушарий преимущественно играет афферентную творческую роль. Наряду с этим она выполняет и моторные функции.
3.15.1. Афферентные функции коры больших полушарий
Афферентные функции коры больших полушарий определяются как афферентными влияниями, поступающими в специфические проекционные (зрительную, обонятельную, тактильную, слуховую) зоны по лемнисковым путям, так и восходящими активирующими влияниями подкорковых образований на кору головного мозга.
К. Бродман по особенностям цитоархитектонического строения разделил кору больших полушарий на цитоархитектонические поля (рис. 3.35).
Особенности проекционных зон.Общим для проекционных областей коры большого мозга является наличие большого количества специфических нейронов, которые дают реакции на раздражители строго определенной сенсорной модальности. Среди спе-
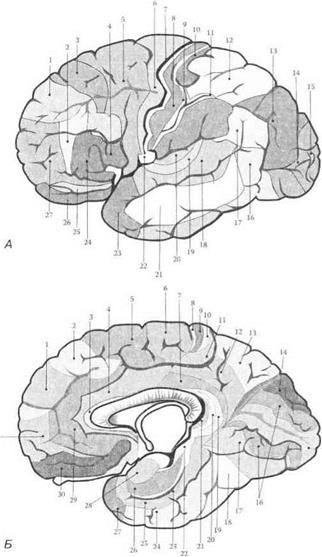
Рис. 3.35.Цитоархитектоническая карта новой коры больших полушарий (по К. Бродману, 1925):
А — латеральная поверхность; Б — медиальная поверхность. Цифры — корковые поля
цифических нейронов выделены проекционные нейроны, имеющие однозначную связь с проекционным рецептивным периферическим полем, и непроекционные нейроны, возбуждающиеся с различных рецептивных полей. Среди нейронов проекционной области коры имеются и неспецифические нейроны, которые реагируют на раздражители различной сенсорной модальности, не относящейся к данной проекционной области. Например, некоторые нейроны зрительной области коры большого мозга реагируют на звуковые стимулы. Неспецифические нейроны, как правило, находятся в ассоциативных областях коры головного мозга.
Специфические проекционные зоны коры больших полушарий. Взадней центральной извилине представлена кожно-соматическая чувствительность: в сенсомоторной коре — тактильная, температурная и болевая, в зрительной коре — световая, в височной коре — звуковая, в инсулярной коре — вкусовая, тактильная, звуковая и световая.
Афферентные проекции в кору больших полушарий.Кора постцентральной извилины рассматривается как первая соматосенсорная область. Сюда проецируется кожная чувствительность противоположной стороны тела от тактильных, болевых, температурных, вкусовых рецепторов, интероцепторов и проприоцепторов в виде «сенсорного гомункулуса» (рис. 3.36). Вторая соматосенсорная кора расположена на границе пересечения центральной борозды с верхним краем височной доли в глубине латеральной борозды. Сигналы в эту область поступают с обеих сторон тела и от других областей мозга, в частности — слуховой и зрительной.
Предполагают участие этих зон коры в контроле движений.
В специфических проекционных зонах наблюдается сенсорнобиологическая конвергенция возбуждений, когда к одним и тем же нейронам поступают сенсорные возбуждения и возбуждения от мотивациогенных центров гипоталамуса и лимбических структур головного мозга.
3.15.2. Эфферентные функции коры больших полушарий
Предцентральная извилина (поле 4) рассматривается как первичная двигательная кора. Вней представлена четкая топографическая
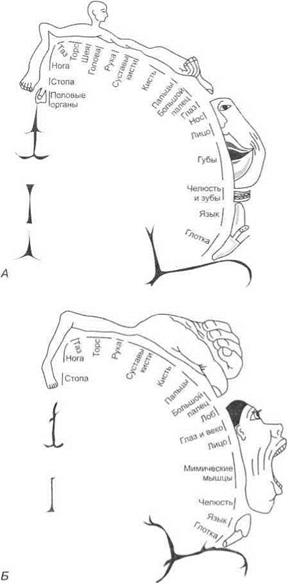
Рис. 3.36.Сенсорные {А) и моторные (Б) представительства в коре больших полушарий
проекция мышц противоположной стороны тела — «двигательный человечек Пенфилда» (см. рис. 3.36). При поражении этой корковой зоны утрачивается способность к тонким координированным движениям, особенно пальцев рук.
Вторичная двигательная кора(поле 6) расположена на латеральной поверхности полушарий, впереди предцентральной извилины (премоторная кора) и на медиальной поверхности верхней лобной извилины. Эта зона коры мозга связана с планированием и координацией произвольных движений.
Вторичная двигательная область дает начало пирамидному тракту и некоторым нисходящим путям экстрапирамидной системы. На пирамидные клетки моторной коры широко конвергируют возбуждения различной сенсорной и биологической модальностей.
У человека в заднем отделе нижней лобной извилины левого полушария (поле 44) расположен центр моторной речи Брока, обеспечивающий речевой праксис (целенаправленная речь), а также музыкальный моторный центр и центр тональности речи (поле 45). Нарушение этого центра приводит к поражению речи.
Возбуждение от пирамидных нейронов по пирамидным трактам распространяется к мотонейронам передних рогов спинного мозга, определяя фазические и тонические воздействия на различные мышечные группы. Одновременно копии эффекторных команд по коллатералям пирамидных трактов распространяются к вставочным нейронам различных отделов коры и подкорковых образований. Именно эти нейроны с помощью поступающей к ним обратной афферентации оценивают эффективность достижения субъектами результатов их деятельности, удовлетворяющих различные потребности.
3.15.3. Ассоциативные зоны
Среди ассоциативных областей коры большого мозга префронтальные области (лобные доли) играют главную роль в выработке стратегии поведения. Выбор стратегии поведения отчетливо проявляется в ситуациях, когда необходим быстрый переход от одних поведенческих актов к другим. Исследования активности нейронов префронтальной области коры в процессе осуществления целенаправленных поведенческих актов показали, что изменения их импульсной активности связаны, главным образом, с моментом выбора цели, механизмами внимания и кратковременной памяти. Передний отдел лобной доли принимает также участие в формировании личностных качеств и творческих процессов человека.
В затылочно-теменной ассоциативной области коры большого мозга осуществляются высший анализ и интеграция соматосенсорных, вкусовых и зрительных сигналов. На нейронах этой области за счет механизма конвергенции объединяются возбуждения, отражающие физические параметры раздражителей, например, массы, размеров, цвета и др. В этих зонах коры наблюдается перекрытие различных видов чувствительности.
Ассоциативной является также теменная область коры. Она участвует в формировании субъективного представления об окружающем пространстве и теле человека.
На нейроны этой ассоциативной области коры головного мозга конвергируют возбуждения различной сенсорной модальности — мультисенсорная конвергенция.
Благодаря генерализованным восходящим активирующим влияниям на многие нейроны различных отделов ассоциативной коры головного мозга конвергируются возбуждения различной биологической модальности — мультибиологическая конвергенция.
Обширные зоны ассоциативной височной доли коры мозга включаются в механизмы долговременной памяти.
Оценка достигнутых результатов в ассоциативных зонах коры осуществляется специальным видом конвергенции возбуждений на нейронах — так называемая аксонально-сенсорно-биологической конвергенция.При этом к одним и тем же нейронам, особенно лобных долей коры мозга, распространяются возбуждения сенсорной и биологической модальностей, а также от коллатералей пирамидного тракта (см. разд. 8.4).
В коре заднего отдела верхней височной извилины левого полушария расположен слуховой центр речи (центр Вернике), поражение которого приводит к нарушению понимания устной речи.
Локализация функций в коре больших полушарий.Кора головного мозга подобна большому телевизионному экрану, на который из внутреннего и внешнего мира постоянно проецируются возбуждения сенсорной и биологической модальности, а также обратная афферентация о результатах совершенных субъектом дей-
| Рис. 3.37.Локализация функций в коре большого мозга человека (по Экономо и Коскинас) |
| Глава 3. Нервная система |
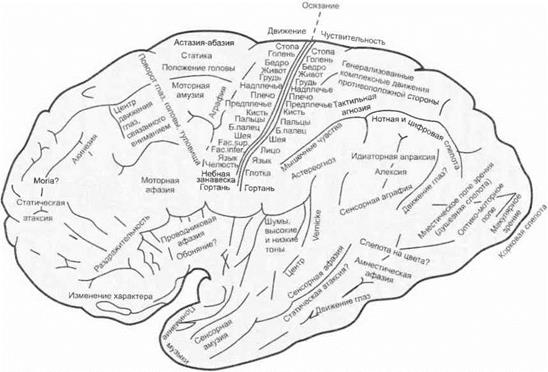
ствий. Распределение этих проекций по коре мозга неоднородно. К одним и тем же нейронам коры мозга поступают как возбуждения по специфическим (лемнисковым) путям, так и восходящие активирующие влияния подкорковых образований. Локализация функций в коре больших полушарий представлена на рис. 3.37.
В настоящее время нельзя говорить ни об узкой локализации корковых центров, ни об их эквипотенциальности (равнозначности). Благодаря тесному взаимодействию возбуждений в коре мозга по вертикальному и горизонтальному типу можно говорить о динамической локализации функций, что определяет процессы компенсации нарушенных функций коры больших полушарий при ее повреждении.
3.15.4. Функции левого и правого полушарий головного мозга
Два полушария большого мозга объединяются мозолистым телом, волокна которого связывают идентичные пункты коры большого мозга и обеспечивают единство ее функционирования. При перерезке мозолистого тела каждое полушарие начинает функционировать независимо одно от другого.
Объективно межполушарную функциональную асимметрию можно выявить по изменениям ЭЭГ на основе сопоставления амплитуды и ритма левого и правого полушарий. У большинства праворуких людей доминирующим является левое полушарие, которое обеспечивает функцию речи. Как указывалось выше, в лобно-височном отделе левого полушария (поле 6а) у человека располагается центр речи Брока. Кроме того, левое полушарие осуществляет контроль за действием правой руки, а также вербальное и логическое мышление.
Правое полушарие головного мозга специализировано для восприятия формы и пространства и участвует в интуитивном мышлении, а также в формировании эмоций.
Доминирование правого полушария проявляется у человека в конкретных видах деятельности, в способности тонко чувствовать и переживать.
В системной организации функций оба полушария взаимосо- действуют достижению полезных приспособительных результатов различных функциональных систем.
3.16. ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ ГОЛОВНОГО МОЗГА
Выше рассмотрены основные физиологические функции разных отделов ЦНС. Однако надо иметь в виду, что деятельность того или иного участка мозга осуществляется в целом организме в неразрывной связи его с другими отделами мозга. Поэтому всегда следует ставить вопрос: что вносит та или иная структура мозга в его целостную деятельность?
Как целостное образование головной мозг постоянно функционирует на основе следующих ведущих механизмов:
• восходящие активирующие влияния подкорковых образований на кору головного мозга;
• нисходящие (активирующие и тормозящие) влияния корковых нейронов на подкорковые образования;
• корково-подкорковая реверберация возбуждений;
• постоянная оценка параметров достигнутых результатов с помощью обратной афферентации;
• механизмы доминанты;
• правое и левое полушария взаимно дополняют друг друга.
В функциональные системы организма, обеспечивающие своей саморегуляторной деятельностью различные полезные для организма показатели, мозговые структуры различного уровня включаются строго избирательно. Это касается синаптических образований нейронов, а также постсинаптических реакций нейронов мозга.
Мозг как целостное образование объединяет центральные механизмы множества функциональных систем гомеостатического, поведенческого и психического уровней.
Ведущим принципом системной организации мозга является принцип доминанты. Во всех случаях деятельность мозга определяет доминирующая по биологической и социальной значимости и формирующаяся на ее основе мотивация, которая определяет выживаемость или адаптацию организма к условиям существования. После удовлетворения доминирующей потребности начинает доминировать следующая потребность.
Системный принцип организации деятельности мозга указывает на то, что распространенная линейная проводниковая парадигма его организации хотя и имеет место, но утрачивает свое универсальное значение.
Интегрированные свойства мозга проявляются также в его химической организации.
3.16.1. Химическая интеграция функций
головного мозга
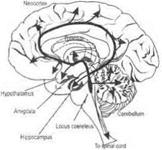
Норадренергические механизмы.Норадренергические нейроны располагаются в голубоватом ядре и прилежащих участках продолговатого и среднего мозга (вентролатеральные ядра ретикулярной формации). Аксоны этих нейронов широко распространяются в стволе мозга, мозжечке, гипоталамусе, гиппокампе, миндалине, поясной и энторинальной коре (рис. 3.38, А).
 | |||||
 | |||||
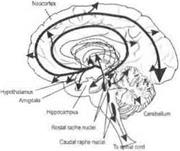 | |||||
|
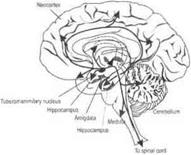 |
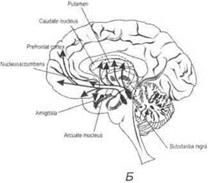
Дофаминергические механизмы.Основные нейроны располагаются в среднем мозге и гипоталамусе.
Одни дофаминергические нейроны, располагающиеся в гипоталамусе, посылают короткие аксоны в гипофиз. Другие дофаминергические нейроны располагаются в черной субстанции. Аксоны этих нейронов проецируются в полосатые тела. Тела третьих дофаминергических нейронов локализуются в среднем мозге рядом с черной субстанцией. Их аксоны проецируются в лимбические структуры (миндалину, обонятельный бугорок, септальную область, поясную извилину и энторинальную кору) и кору головного мозга, особенно ее фронтальные отделы (рис. 3.38, Б).
Серотонинергические механизмы.Серотонинергические нейроны локализуются в дорсальном и медиальном ядрах шва продолговатого мозга, а также в среднем мозге и варолиевом мосту. Их аксоны иннервируют гипоталамус, миндалины, бледный шар, гиппокамп и кору больших полушарий. Высокая концентрация серотонина обнаружена в эпифизе (рис. 3.38, В).
Холинергические механизмы.Тела холинергических нейронов находятся в ядрах перегородки, диагонального пучка (Брока) и в базальных ядрах. Их аксоны проецируются к структурам переднего мозга, особенно в гиппокамп и новую кору (рис. 3.38, Г).
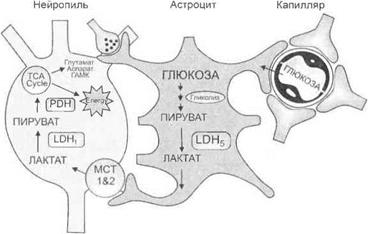
Рис. 3.39.Нейрохимическое взаимодействие астроцита с синапсом
Медиаторные функции олигопептидов и аминокислот.Оли- гопептиды-опиоиды, вещество П, ангиотензин II, вазопрессин, окситоцин и др., а также цитокины — модулируют действие медиаторов в синапсах ЦНС. Самостоятельную медиаторную функцию выполняют аминокислоты — глутаминовая кислота, аспарагиновая кислота и ГАМК. У ряда нейронов ЦНС выявлены специфические рецепторы к глутамату (NMDA-рецепторы, аффинные к N-метил- D-аспартату).
ГАМК — самый распространенный тормозной медиатор в ЦНС. Другим тормозным медиатором, точкой приложения которого являются нейроны продолговатого и спинного мозга, является глицин.
Взаимодействие нервных и глиальных элементов.В интегративной деятельности ЦНС осуществляется тесное взаимодействие нейронов с окружающими их глиальными клетками и кровеносными сосудами. На рис. 3.39 схематически показаны механизмы утилизации глюкозы, поступающей из мозгового капилляра в астроцит, ее переработка в лактат и глутамат с участием АТФ. Глутамат в дальнейшем участвует в механизме синаптической передачи.
3.17. ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА
Вегетативная (автономная) нервная система — часть общей нервной системы, имеющая отношение преимущественно к регуляции вегетативных (растительных) функций: гладкой мускулатуры, железистого аппарата, соединительной ткани внутренних органов, кровеносных и лимфатических сосудов. Вегетативная нервная система отчасти оказывает влияние на деятельность скелетной мускулатуры и наружных половых органов. Вегетативная нервная система рассматривается как автономная, так как ее деятельность в слабой степени регулируется произвольно.
Выделение вегетативной (автономной) нервной системы условно. В целостных системных реакциях организма вегетативные функции всегда тесно взаимодействуют с соматическими (см. гл. 6). Все это заставляет говорить о вегетативных компонентах целостных приспособительных реакций.
Морфофункциональная организация вегетативной нервной системы.На основании структурно-функциональных особенностей
различают три отдела вегетативной нервной системы: симпатический, парасимпатический и метасимпатический.
Сравнительная характеристика этих отделов, а также соматической нервной системы представлена в табл. 3.1.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|