- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Вегетативная нервная система 1 страница
Глава 3
Нервная система
Морфофункционально нервная система подразделяется на центральную, периферическую, соматическую и вегетативную.
Нервная система определяет восприятие метаболических потребностей, оценку состояния различных органов и тканей организма, воздействий внешней и внутренней среды и регуляцию функций организма.
3.1. РЕЦЕПТОРЫ НЕРВОВ
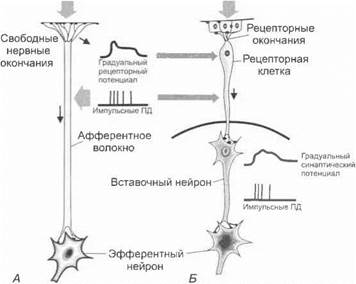
Рецепторы окончаний нервов.Рецепторы нервов представлены свободными немиелизированными нервными окончаниями или специальными рецепторными органами (см. разд. 1.7). В первич- ночувствующих рецепторах возбуждение формируется непосредственно при действии раздражителя на мембраны нервных окончаний. Во вторичночувствующих рецепторах первично возбуждаются ненервные рецепторные клетки (рис. 3.1).
3.2. ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА НЕРВОВ
Нерв как ткань обладает свойством возбудимости и проводимости. В ответ на раздражение в нервах возникают биопотенциалы (см. разд. 1.3).
|
|
 |
Возбужденный участок нерва становится электроотрицательным по отношению к соседним участкам. Благодаря разности потенциалов между возбужденным и невозбужденным участком нерва процесс возбуждения распространяется вдоль нервного волокна.
Бездекрементное (незатухающее) распространение возбуждения по нервам осуществляется за счет использования энергии самого нервного субстрата, в частности АТФ.
Соматические нервные волокна по сравнению с мышцами и нервно-мышечными синапсами обладают высокой функциональной лабильностью.
Особенности проведения возбуждения по нервам
Закон двустороннего проведения возбуждения по нерву.При нанесении раздражения на нерв возбуждение по нерву без затухания распространяется в обе стороны. Это можно зарегистрировать при отведении биопотенциалов с двух сторон от пункта раздражения.
Закон изолированного проведения возбуждения в отдельных нервных волокнах внутри нервного ствола.Каждое нервное волокно надежно изолировано миелиновой оболочкой.
Закон физиологической и морфологической целостности нерва.Возбуждение не распространяется через поврежденный нерв. Температурные воздействия, воздействие анодом постоянного тока и действие химических веществ блокируют проведение возбуждения по нерву. Особенно значимо для проведения возбуждения по нерву его кровоснабжение. Прекращение кровоснабжения нарушает проведение возбуждения по нерву.
Относительная неутомляемость нерва.При наложении на нерв нервно-мышечного препарата льда или при действии анода постоянного тока при удалении этих блокирующих воздействий можно наблюдать эффекты мышечных сокращений после многочасового непрерывного электрического раздражения нерва выше участка альтерации.
Низкое теплообразование в нерве:7—10-8 кал при одиночном возбуждении.
Обе фазы теплообразования в нерве — аэробные. В нерве имеются специальные донаторы кислорода.
Возбуждение распространяется по нерву двумя способами:линейное — вдоль его оси и сальтаторное (скачкообразное) — через перехваты Ранвье. В перехватах Ранвье нервные волокна не покрыты миелином. Электрополевые воздействия, связанные с процессом возбуждения, оказывают раздражающее воздействие на 2—3 соседних перехвата Ранвье. Благодаря этому обеспечивается высокая скорость распространения возбуждения по нерву и перестраховываются локальные повреждения нервных волокон.
Сальтаторный тип распространения возбуждения присущ только миелиновым волокнам. Этот механизм не имеет места у вегетативных нервов, не покрытых миелиновой оболочкой.
Соматические нервы характеризуются высокой скоростью проведения возбуждения. У лягушки — 20—30 м/с, у теплокровных — 100-160 м/с.
Различные нервные волокна характеризуются различной возбудимостью и скоростью проведения возбуждения.
В 1936 г. Гассер и Эрлангер нанесли раздражение на участок нервного ствола и зарегистрировали биопотенциалы на разных расстояниях от места раздражения. При этом в разных участках нерва потенциалы обнаружили различную форму (рис. 3.2).
Опыт свидетельствовал о том, что нервный ствол неоднороден. Разные составляющие его волокна имеют различную скорость проведения возбуждения.
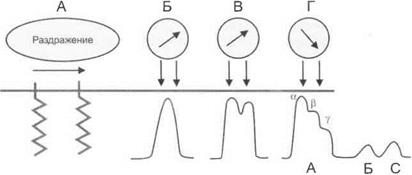
Рис. 3.2.При регистрации процесса возбуждения на разном удалении от пункта раздражения (А), в точках Б, В, Г потенциалы действия изменяются. В точке Г регистрируются волны А, Б, С и а, Р, у, отражающие различную скорость проведения возбуждения в составляющих нервный ствол волокнах
Волокна А (а, р, у) — толстые миелиновые волокна, диаметром более 15 мкм. Скорость проведения возбуждения по волокнам Аа у человека — 100—160 м/с, Ар — 40 м/с, Ау — 10 м/с.
Волокна В — тонкие миелинизированные (например, преган- глионарные вегетативные волокна), диаметром — 10—11 мкм. Скорость проведения возбуждения по волокнам В — менее 10 м/с.
Волокна С — тонкие безмиелиновые (например, постганглионарные волокна вегетативной нервной системы, болевые волокна), диаметром 4—5 мкм. Скорость проведения возбуждения по волокнам С — 0,7 м/с.
В нервных стволах возбуждение одних волокон влияет на состояние других (эфаптическое действие). При этом через воздействия на перехваты Ранвье в соседних волокнах возникают локальные потенциалы, которые могут суммироваться и вызывать в них распространяющийся процесс возбуждения.
3.3. ФИЗИОЛОГИЯ СИНАПСОВ
Среди других разнообразных тканей организма особая роль принадлежит синаптической области. С помощью синапсов осуществляется взаимодействие разнородных по функциям тканей организма. Это взаимодействие происходит на основе «химического компромисса». По сравнению с взаимодействующими тканями синаптическая область характеризуется специфическими химическими свойствами. Это впервые было показано классическим опытом французского физиолога Клода Бернара.
Опыт состоял в том, что в чашку Петри, заполненную стрельным ядом индейцев кураре[I], помещали два нервно-мышечных препарата. Один препарат (1) располагали таким образом, что в раствор кураре погружался только нерв. Другой препарат (2) размещали таким образом, что в раствор кураре попадали мышца, нерв и участок соединения нерва с мышцей. На нервы и мышцы обоих препаратов устанавливали электроды, с помощью которых осуществляли электрическое раздражение этих тканей.
При непрямом (через нерв) электрическом раздражении нервно-мышечного препарата 1, несмотря на то, что его нерв долго располагался в растворе кураре, мышца этого препарата отвечала сокращениями. На прямое электрическое раздражение эта мышца тоже реагировала сокращением. Иная картина наблюдалась у нервно-мышечного препарата 2. На непрямое электрическое раздражение нерва нервно-мышечного препарата 2 мышца этого препарата не сокращалась. Однако на прямое электрическое раздражение этой мышцы наблюдалось ее сокращение. Вывод: кураре не действует на нерв и мышцу, а действует на место их соединения, которое, по сравнению с нервом и мышцей, обладает особыми химическими свойствами.
Строение синаптической области.Термин «синапс» (от греч. synapsis — контакт, застежка) введен в физиологию английским физиологом Ч. Шеррингтоном, исходя из того, что под световым микроскопом на телах нервных клеток контакты аксонов выглядят как «застежки». Научные представления о строении синапсов существенно продвинулись в связи с открытием электронного микроскопа. Электронно-микроскопические исследования позволили установить, что синаптическая область имеет сложное строение (рис. 3.3). В ней различают пресинаптическую область, включающую окончание приходящего аксона, — синаптическая колба. Характерно, что в пресинаптической области миелинизированные нервы теряют миелиновую оболочку. В пресинаптической области выделяют пресинаптическую мембрану — более утолщенный участок мембраны синаптической колбы, прилежащей к иннервируемой ткани. Внутри синаптической колбы находятся синаптические пузырьки и митохондрии. Синаптические пузырьки заполнены медиатором. Функция митохондрий связана с энергетическим обменом в пресинаптической области.
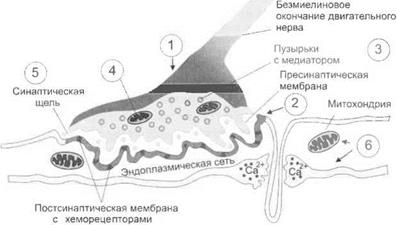
Рис. 3.3.Морфофункциональная организация синаптической области: 7 — пресинаптическое нервное окончание; 2— постсинаптическая область; 3— везикулы, содержащие медиатор; 4— митохондрии; 5— синаптическая щель; 6— постсинаптические органеллы
Противолежащий пресинаптической мембране участок мембраны — постсинаптическая мембрана. Этот участок мембраны тоже более утолщен по сравнению с другими участками мембраны иннервируемой ткани. Постсинаптическая мембрана отделяет от пресинаптической постсинаптическую область — клеточную структуру со всеми типичными для нее ингредиентами, включая ядро клетки. От постсинаптической мембраны к ядру протягиваются нежные мо
лекулярные трубочки, образованные молекулами специфических белков.
Пресинаптическая область отделяется от постсинаптической так называемой синаптической щелью порядка 200—1000 А (ангстрем — одна десятитысячная микрона). Синаптическая щель заполнена межтканевой жидкостью с ее ингредиентами.
Организация синапсов. Ворганизации синаптической области существенен вопрос: чему в морфофункциональном отношении принадлежит синапс? Принадлежит ли он нервной ткани или ткани постсинаптической области?
Ответ на этот вопрос дают следующие эксперименты.
При перерезке подходящего к синапсу нерва синапсы в соответствующих тканях, как показывают гистологические исследования, исчезают. После сшивания и реиннервации синаптической области синапсы снова восстанавливаются. Таким образом, теоретически синапс организуется иннервируемым нервом. В настоящее время выявлены специальные белковые вещества, которые вырабатываются в нервной ткани и способствуют организации синапсов — си- наптины.
Организации синапсов способствуют также факторы роста нервов и инсулин. Однако морфологическое восстановление синапсов после реиннервации еще не говорит о восстановлении их функции.
Эксперименты с гетерогенными анастомозами нервов, когда сшивается центральный конец одного нерва с периферическим концом чужеродного для него нерва, выявили еще одну особенность организации синаптической области. При сшивании даже разнородных нервов нейрофибриллы из центрального конца сшитых нервов начинают прорастать в периферический ствол чужеродного нерва со скоростью 6—8 мм/сут и, в конечном счете, устанавливают синаптические контакты с чужеродной для них тканью. Морфологически при этом синапсы также восстанавливаются полностью. Однако в функциональном отношении выявляются следующие закономерности, которые лучше всего проследить на примере гетерогенного анастомоза центрального конца диафрагмального нерва (n. Frenicus) с периферическим концом перерезанного нерва барабанной струны (n. Chorda Tympani), иннервирующего обычно подчелюстную слюнную железу.
В результате анастомоза диафрагмального нерва с нервом барабанной струны эфферентные волокна, находящиеся в составе центрального отрезка ствола диафрагмального нерва, прорастают в периферический конец перерезанного нерва барабанной струны и восстанавливают синапсы в ткани подчелюстной слюнной железы. В остром опыте электрическое раздражение диафрагмального нерва выше и ниже места анастомоза электрическими импульсами низкой частоты (до 10 имп./с) вызывало слюноотделение подчелюстной слюнной железой. Это указывало на проходимость импульсов, идущих по волокнам диафрагмального нерва синаптической области в подчелюстной слюнной железе.
Если же применяли высокочастотное раздражение (свыше 10 имп./с), раздражение диафрагмального нерва не вызывало слюноотделения. Значит, восстановленные синапсы пропускали низкочастотные импульсы и не пропускали высокочастотные. Как указывалось ранее, в естественных условиях по диафрагмальным нервам в организме распространяется высокочастотная импульсация до 1000 имп./с.
Такая высокочастотная импульсация не проходит через вновь образованные на слюнной железе синапсы. Из этого опыта можно сделать вывод: несмотря на то, что структурный синапс организуется приходящим к тканям нервом, функциональные свойства, и в частности способность проводить возбуждения, определяются самой тканью, свойствами постсинаптической области (и в первую очередь — ее функциональной лабильностью).
Таким образом, синапс структурно создается нервом, а его физиологические свойства зависят от свойств иннервируемой ткани, и постсинаптической мембраны в частности.
Механизм проведения возбуждения через синапс.Структурные и функциональные свойства синапсов определяют механизм распространения возбуждения через синаптическую область.
Классификация синапсов.С морфологической точки зрения в зависимости от свойств объединяемых тканей синапсы подразделяются на нервно-мышечные, нервно-секреторные и межнейрональные. Нервно-мышечные синапсы, в отличие от других, имеют особенности морфологической организации. Они состоят в том, что пресинаптический участок представлен не в виде колбы, а широко пилообразно разветвлен между мышечными волокнами (см. рис. 3.3). С функциональной точки зрения синапсы подразделяются на возбуждающие (проводящие возбуждение) и тормозные (блокирующие проведение возбуждения).
В зависимости от характера выделяющегося в синапсах медиатора синапсы подразделяют на адренергические, холинергические, серотонинергические, ГАМКергические и др.
Активность синапсов в состоянии покоя.В состоянии покоя на постсинаптической мембране с помощью подведенных к ней микроэлектродов можно зарегистрировать периодически возникающие слабые процессы деполяризации, так называемые «миниатюрные потенциалы». Напряжение этих потенциалов — порядка 0,5 мВ. Происхождение «миниатюрных потенциалов» связывают с периодическим опорожнением медиаторов из отдельных везикул пресинаптической области на постсинаптическую мембрану. Мембраны везикул обладают протеолитической активностью. Как только везикула соприкасается с пресинаптической мембраной, происходит лизис везикулярной и пресинаптической мембран — и содержимое везикулы изливается на постсинаптическую мембрану, вызывая ее деполяризацию, типа местного потенциала. При этом медиатор изменяет проницаемость ионных каналов постсинаптической мембраны. Ионы Na+ устремляются с поверхности через постсинаптическую мембрану внутрь протоплазмы клеток постсинаптической ткани. Ионы К+, наоборот, устремляются на поверхность постсинаптической мембраны.
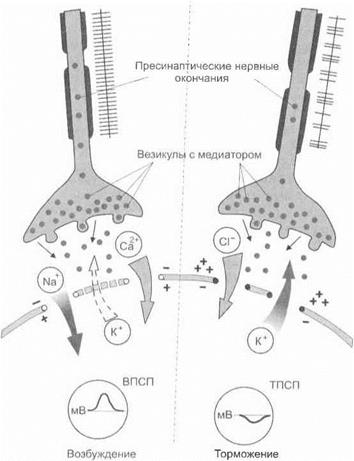
Активность синапсов в процессе распространяющегося возбуждения.Химическая теория. При поступлении к пресинаптической области распространяющегося процесса возбуждения в пресинаптической мембране открываются потенциалзависимые кальциевые каналы и ионы Са2+ устремляются в пресинаптиче- скую колбу. Под влиянием ионов Са2+ пресинаптические везикулы начинают массивно концентрироваться около пресинаптической мембраны. При этом происходит массивный лизис пресинаптической мембраны и большие количества медиатора поступают на постсинаптическую мембрану. В случае, если везикулы содержат возбуждающий медиатор, это приводит к обширной деполяризации постсинаптической мембраны, суммации на ней местных потенциалов и возникновению распространяющегося спайкового процесса возбуждения. На постсинаптической мембране с помощью микроэлектрода в этом случае регистрируются возбуждающие постсинаптические потенциалы (ВПСП). В случае, если пресинаптические везикулы синапса содержат тормозной медиатор, например ГАМК, при его поступлении к постсинаптической мембране происходит ее гиперполяризация и регистрируется тормозной постсинаптический потенциал.
Природа постсинаптического возбуждения представляется следующим образом. При поступлении возбуждающего медиатора (например, ацетилхолина) к постсинаптической мембране активируются ее натриевые каналы. При этом ионы Na+ устремляются внутрь постсинаптической клетки, а ионы К+ — наружу. Происходит деполяризация мембраны (рис. 3.4). В свою очередь ацетилхолин активно разрушается в постсинаптической области с помо
рие. 3.4.Молекулярные механизмы в возбуждающем и тормозном синапсах
щью фермента холинэстеразы и распадается на холин и уксусную кислоту. В результате этого процесс возбуждения прекращается и возникает снова при поступлении следующей достаточной порции ацетилхолина. Деполяризации способствуют ионы Са2+, освобождающиеся из пресинаптической области. Возбуждение постсинаптической мембраны носит, таким образом, импульсный, дискретный характер. Порция медиатора, достаточная для того, чтобы вызвать процесс деполяризации постсинаптической мембраны, получила название «кванта» медиатора.
В возбуждении постсинаптической мембраны принимают участие и другие выделяющиеся в пресинаптической области вещества: ионы Са2+, отдельные олигопептиды, такие как вещество П, факторы роста нервов и т.д.
В тормозных синапсах поступающий на постсинаптическую мембрану тормозной медиатор активирует выход ионов К+ из клетки, блокирует натриевые каналы и создает процесс гиперполяризации мембраны. При этом в постсинаптическую область проникают ионы С1".
Рассмотренные механизмы составляют сущность химической теории возбуждения синапсов, основы которой заложены русским физиологом А. Ф. Самойловым и американским исследователем, лауреатом Нобелевской премии Дж. Экклсом.
В настоящее время химическая теория возбуждения синаптической области значительно расширилась. В 1970-е гг. П. К. Анохин первый указал на ограниченность мембранной теории возбуждения и призвал исследовать постсинаптические процессы возбуждения.
Постсинаптические процессы возбуждения.Постсинаптические процессы представляются следующим образом. Медиаторы на постсинаптических мембранах активируют специальные белковые молекулы-рецепторы. Эти рецепторы в свою очередь активируют специальные G-белки. Последние возбуждают фермент аденилат - циклазу. Аденилатциклаза активирует циклические аденозинмонофосфат (цАМФ) и гуанозинмонофосфат (цГМФ). Будучи ферментами, цАМФ и цГМФ фосфорилируют белки. Наряду с этим функцию вторичных передатчиков (мессенджеров) играют инозитолы, ионы Са2+ и различные протеинкиназы.
Образование медиаторов.Специального внимания заслуживают процессы образования медиатора в синаптической области. Согласно одной точке зрения, медиатор вырабатывается на месте в пресинаптических везикулах за счет постоянного их образования из пресинаптических мембран путем пиноцитоза.
Другая точка зрения связывает образование медиатора в теле нейрона, в частности в аппарате Гольджи, с поступлением его по каналам аксона в синаптическую область. Подтверждением этой точки зрения являются опыты с перевязкой аксонов. В этом случае содержание медиаторов в соответствующих синаптических областях существенно снижается.
Электрическая теория возбуждения синапсов.Наряду с химической существует электрическая теория передачи возбуждения через синапсы. При этом в качестве причин деполяризации постсинаптической мембраны рассматривают влияние электромагнитных полей, которые создаются в пресинаптической области при поступлении к пресинаптической области импульсов возбуждения.
Возникающие вокруг пресинаптической области электрополе- вые влияния распространяются по жидким средам до постсинаптической мембраны и посредством суммации местных процессов возбуждения могут вызвать процесс распространяющейся деполяризации. Возбуждающиеся на электрической основе синапсы получили название эфапсы. Эфапсы широко представлены у простейших животных (например, моллюсков). Они имеются в ЦНС и у человека, особенно в областях древнего мозга (например, гиппокампа). Не исключено, что через эфапсы осуществляется воздействие на мозг электромагнитных излучений.
В отдельных синапсах в проведении возбуждения принимают участие оба — химический и электрический — механизма. При поступлении к пресинаптической области этих синапсов импульсов возбуждения за счет электрополе вых влияний в короткие сроки активируется постсинаптическая мембрана. Процесс усиливается химическим путем за счет последующего действия синаптических медиаторов.
3.3.1. Общие свойства синапсов
Общие свойства синапсов определяются особенностями их строения и механизмом проведения возбуждения.
Одностороннее проведение возбуждения через синапс.Это свойство определяется особенностями химического механизма проведения возбуждения через синапс. Как было указано ранее, везикулы с медиатором располагаются в пресинаптической области и поступающий в пресинаптическую щель медиатор действует только в одном направлении.
Синаптическая задержка.Возбуждение в синапсах задерживается от 1 до 3 мс. Синаптическая задержка определяется перемещением пресинаптических везикул к пресинаптической мембране, процессами лизиса пресинаптической мембраны, поступлением медиатора в синаптическую щель и действием его на проницаемость ионных каналов постсинаптической мембраны.
Суммация возбуждения в синапсе.Определяется процессами перехода местного процесса возбуждения на постсинаптической мембране в спайковый распространяющийся процесс возбуждения.
Синаптическая область обладает свойством трансформации приходящих к ней возбуждений. Это определяется следующими свойствами синапсов.
Свойство облегчения проведения возбуждения.Это свойство состоит в том, что каждый последующий импульс возбуждения проводится через синапс более облегченно благодаря предшествующим процессам возбуждения.
Свойство проторения проведения возбуждения через синапс.Это свойство означает, что каждый предшествующий импульс возбуждения создает для последующих более облегченные условия формирования.
Свойства облегчения и проторения возбуждения через синапс лежат в основе так называемой синаптической памяти. Синаптическая память связана с формированием в синапсах в процессе их возбуждения специальных белков памяти.
Синапсы обладают более низкой функциональной лабильностьюпо сравнению с нервной, мышечной и даже железистой тканью. Соматические нервы, иннервирующие поперечно-полосатые мышцы, способны проводить без трансформации в среднем до 500 имп./с. Наиболее высоколабильными в организме являются слуховые и диафрагмальные нервы. Их функциональная лабильность превышает 1000 имп./с. Поперечно-полосатые мышцы проводят без трансформации до 200—300 имп./с. Нервно-мышечный синапс обладает функциональной лабильностью до 70 имп./с. Отсюда понятно, что нервно-мышечный, так же как и нервно-секреторный, синапс не способен проводить без трансформации высокую частоту возбуждений.
Повышенная утомляемость синапсов.Синапсы обладают повышенной утомляемостью по сравнению с организующими их тканями. Это свойство синапсов отчетливо демонстрирует опыт на нервно-мышечном препарате. При ритмическом непрямом электрическом раздражении нервно-мышечного препарата можно через некоторое время наблюдать утомление мышцы. Однако при прямом дополнительном электрическом раздражении мышцы она демонстрирует отчетливые сокращения. Нерв, как указывалось выше, практически не утомляем. Таким образом, утомление нервно-мышечного препарата при непрямом раздражении связано с первичным утомлением нервно-мышечного синапса.
Синапсы обладают повышенной и избирательной чувствительностью по отношению к химическим и фармакологическим веществам и ядампо сравнению с действием этих веществ на составляющие их ткани. Примером избирательного действия на нервно-мышечный синапс является действие кураре. В настоящее время установлено, что кураре избирательно действует на постсинаптическую мембрану поперечно-полосатой мышцы и блокирует на ней процесс перехода местного потенциала в распространяющийся.
Установлены химические вещества, избирательно действующие на химические процессы распространения возбуждения от постсинаптической мембраны к ядру клеток постсинаптической области, а также на процессы синтеза белка геномом ядер этих клеток.
Повышение чувствительности денервированных структур.Как уже указывалось выше, организующие синапсы нервы обладают трофической функцией. После денервации изменяется чувствительность мембран тканей постсинаптической области к медиаторам. При этом не только мембрана постсинаптической, но и прилежащих областей приобретает способность реагировать на медиатор.
Синапсы — места регулирования процесса возбуждения.Приведенные факты свидетельствуют о том, что на уровне синаптических образований процесс возбуждения приобретает свойства регулируемости. В синапсах процесс возбуждения может быть уси
лен или заблокирован и даже качественно изменен. По аналогии с техническими электронными устройствами синапсы выступают в роли универсальных усилителей, трансформаторов и выпрямителей. Синапсы в процессах жизнедеятельности играют исключительно важную роль, поскольку действие фармакологических веществ в первую очередь направлено на специфические синаптические образования, знать которые в каждом случае обязан врач.
Исключительная роль синапсов в процессах жизнедеятельности в последние годы позволила выделить самостоятельный раздел знаний — синаптологию — как науку, изучающую различные свойства синапсов в организме и их реакции на разнообразные воздействия.
3.4. ФИЗИОЛОГИЯ НЕЙРОНОВ
Нейрон — структурная и функциональная единица центральной нервной системы.
Сформулированы следующие принципы нейронной организации (Р. Кахаль):
• Нейрон с отростками составляет единое морфологическое целое.
• Нейроны генетически едины, происходят из одних нейробластов.
• Нейрон един функционально.
• Возбуждение по нейрону распространяется в одном направлении — от дендритов к аксону.
• Нейрон представляет трофическое единство. При удалении тел нейронов наблюдается гибель аксонов. Перевязка аксона также вызывает гибель нейронов.
• Нейрон участвует как единое целое в патологических реакциях. Деструктивные изменения охватывают весь нейрон.
Нейронная теория.Указанные принципы легли в основу нейронной теории построения ЦНС. Согласно этой теории, строение и функции ЦНС определяются множеством взаимосвязанных нейронов с дискретными свойствами.
Наряду с этим не утрачивают своего значения представления о том, что ЦНС имеет сплошное строение типа синцития, присущее, например, нейропилю, имеющемуся в нервной системе низ
ших беспозвоночных (кишечнополостные) и в мозге земноводных (аксолотли).
Гетерогенные нейроны в их взаимодействиях позволяют ЦНСчеловека участвовать в миллионах разнообразных реакций. На телах отдельных нейронов располагается от сотен до тысяч синапсов, причем отдельные синапсы функционируют с помощью различных медиаторов и связаны со специфическими постсинаптическими химическими реакциями.
Процессы возбуждения нейронов объясняют электрическая и химическая теории.
Электрическая теория возбуждения нейронов.Исходит из представлений о морфофункциональной неоднородности нейронов. У места отхождения аксонов располагается так называемый «светлый аксонный холмик», или «светлое пятнышко». Этот участок нейрона положительно заряжен по отношению к другим участкам мембраны нейронов. При возбуждении синапсов, располагающихся на дендритах и теле нейронов, увеличивается разность потенциалов между «аксонным холмиком» и остальной частью нейрона. При достижении определенной величины это порождает спайковую активность аксона. Чем круче нарастает разность потенциалов тела нейрона и «аксонного холмика», тем чаше выходная импульсация аксона. Все это порождает определенную конфигурацию импульса- ции аксона (рис. 3.5).
 | |||
 | |||
| ||||
| ||||
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|