- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Вегетативная нервная система 2 страница
Химическая теория возбуждения нейронов.Химическая теория связывает процессы возбуждения нейронов со спецификой выделяемых синапсами нейронов медиаторов, олигопептидов и других биологически активных веществ.
Возбуждающие медиаторы (норадреналин, ацетилхолин, глутамат и др.) деполяризуют постсинаптическую мембрану и формируют возбуждающие постсинаптические потенциалы.
Тормозные медиаторы (например, ГАМК) формируют на постсинаптической мембране тормозные постсинаптические потенциалы (рис. 3.6).
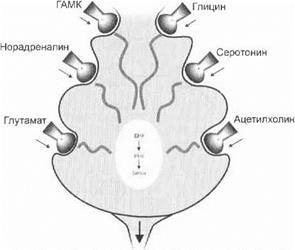
Рис. 3.6.Химическая гетерогенность синаптических образований нейрона
Процесс возбуждения нейронов зависит также от специфики постсинаптических химических реакций, распространяющихся к генетическому аппарату ядер нейронов.
3.4.1. Особенности распространения возбуждения в ЦНС
Распространение возбуждения в ЦНС определяется свойствами внутрицентральных синапсов. Это:
• Одностороннее распространение возбуждения от афферентной части нервной системы к эффекторной.
• Задержка распространения возбуждения в ЦНС (центральная синаптическая задержка). Зависит от числа синапсов, участвующих в реакции.
• Иррадиация возбуждений (рис. 3.7).
Иррадиация может быть системной (направленной) — например, при вытягивании руки возбуждаются только нейроны экстензоров, а флексорные нейроны тормозятся, и диффузной — например, при введении стрихнина.
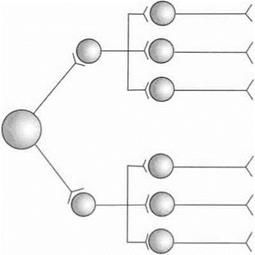
Рис. 3.7.Иррадиация возбуждения в ЦНС
• Временная (последовательная) суммация возбуждений, когда один импульс не проходит через синапс, а серия — проходит.
• Пространственная суммация возбуждений, связанная с конвергенцией поступающих через различные синапсы на одни и те же нейроны различных возбуждений.
• Трансформация возбуждения — увеличение или торможение исходной ритмики возбуждений:
а) способность к облегчению проведения возбуждения. Синапсы ЦНС обладают выраженной экзальтационной фазой изменения возбудимости. У них почти отсутствует рефрактерность. К тому же, в области постсинаптической мембраны при прохождении импульсов возбуждения накапливаются РНК и АТФ, определяющие синаптическую память;
б) способность к проторению возбуждения. Каждый предыдущий импульс прокладывает путь для последующих.
• Умножение (мультипликация, дисперсия) импульсов. За счет разницы во времени синаптических задержек на выходном нейроне происходит умножение числа импульсаций (рис. 3.8).
А , • Ж , ■ Ж ’ лш1
'Ж
2 3
Рис. 3.8.Феномен умножения импульсов в ЦНС. Благодаря синап-
тическим задержкам нейрон 4 получает последовательно три возбуж-
дения
• Пролонгирование (удлинение времени) возбуждения ЦНС. В ЦНС обнаружены специальные «нейронные ловушки» (Ло- ренто Де Но), в которых возбуждение может циркулировать длительное время, пока не будет прекращено тормозным процессом (рис. 3.9).
• Принцип общего конечного пути. Определяется множественной конвергенцией возбуждений на исполнительных нейронах. Ч. Шеррингтон сравнивал конечные исполнительные
Л ,1 Л 2 ААЛ
К Л
Рис. 3.9.«Нейронная ловушка»
нейроны с «воронкой» и рассматривал их как «чек на предъявителя». Как правило, конечными нейронами завладевает более сильное возбуждение.
• Избирательная чувствительность центральных нейронов к действию различных химических веществ. Химические агенты могут действовать на пресинаптическую и постсинаптическую области нейронов мозга, блокируя образование медиаторов, их взаимодействие с постсинаптической мембраной, обратный захват медиатора пресинаптической областью или постсинаптические химические реакции, вплоть до экспрессии генов.
Системная деятельность нейронов.С позиций теории функциональных систем нейроны избирательно, различными синапсами и постсинаптическими процессами включаются в различные функциональные системы и в различные стадии центральной архитектоники функциональных систем (см. разд. 8.4). Этому в значительной степени способствует открытый А. А. Ухтомским принцип доминанты — господствующего возбуждения.
Доминанта в ЦНС.Классический опыт А. А. Ухтомского заключался в электрическом раздражении моторной коры у кошки. Раздражение всегда вызывало сокращение контралатеральной лапы. Однако если в прямую кишку кошки был введен резиновый баллон и его слегка раздували, то то же раздражение моторной коры вместо сокращения лапы вызывало у животного акт дефекации. Аналогично, если в желудок предварительно вводили слабый раствор отвергаемого вещества, при раздражении моторной коры у животных возникала рвота.
Доминирующая потребность, таким образом, создает в ЦНС доминирующее системное возбуждение. Доминирующий очаг возбуждения обладает свойством повышенной возбудимости и притягивает к себе другие возбуждения.
Интегративная деятельность нейронов. П. К.Анохин сформулировал гипотезу, согласно которой процесс возбуждения не ограничивается мембраной нейронов, а распространяется в глубь цитоплазмы постсинаптических неронов. Благодаря множественной конвергенции возбуждений на отдельных нейронах, нейроны, включенные в доминирующее возбуждение, избирательно реагируют только на это возбуждение (принцип ограничения степеней свободы нейрона). При этом каждое из этих возбуждений формирует специфическую постсинаптическую реакцию вплоть до экспрессии геномом нейрона определенного вида белка, формирующего, в свою очередь, специфическую реакцию мембраны нейрона, определяющую его реагирование только на те возбуждения, которые способствуют удовлетворению исходной потребности организма.
3.4.2. Процессы торможения в ЦНС
Наряду с механизмами возбуждения в ЦНС существуют механизмы торможения, которые проявляются в прекращении или уменьшении активности нервных клеток. В отличие от возбуждения торможение — локальный нераспространяющийся процесс, возникающий на клеточной мембране.
Сеченовское торможение.Наличие процесса торможения в ЦНС впервые было показано И. М. Сеченовым в 1862 г. в экспериментах на лягушках.
И. М. Сеченов осуществлял разрез головного мозга лягушки на уровне зрительных бугров и измерял время рефлекса отдергивания задней лапы при погружении ее в раствор соляной кислоты (рефлекс Тюрка). При наложении на разрез зрительных бугров кристаллика калийной соли время рефлекса увеличивалось. Удаление кристаллика приводило к восстановлению исходного времени рефлекторной реакции.
Вначале И. М. Сеченов полагал, что он открыл задерживающие (тормозные) центры в зрительном бугре. Затем его представления
о центральном торможении изменились. И. М. Сеченов пришел к выводу, что торможение является следствием взаимодействия двух и более возбуждений на спинальных нейронах: возбуждения, обусловленного раздражением рецепторов соляной кислотой, и возбуждения, обусловленного раздражением зрительных бугров кристалликом соли. В этом случае одно возбуждение неизбежно становится тормозящим, а другое — тормозимым. По современным представлениям, подавление одним возбуждением другого в опыте И. М. Сеченова
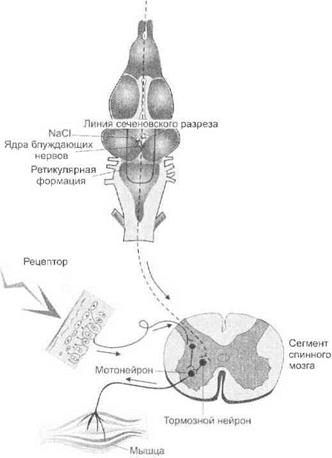
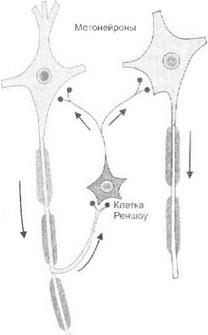
Рис. 3.10.Механизм сеченовского торможения
происходит на уровне сегментов спинного мозга. При этом центральное возбуждение распространяется на вставочные тормозные нейроны (клетки Реншоу), которые оказывают пресинаптическое и постсинаптическое торможение мотонейронов (рис. 3.10).
Пресинаптическое торможение.Пресинаптическое торможение развивается в пресинаптической части синапса за счет воздействия на его мембрану аксо-аксональных синапсов. В результате как деполяризующего, так и гиперполяризующего воздействия происходит блокирование проведения импульсов возбуждения по пресинап- тическим путям к постсинаптической нервной клетке (рис. 3.11).
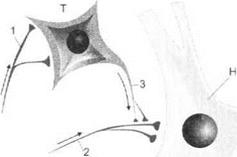

Рис. 3. 11.Пресинаптическое торможение:
Т— тормозной нейрон; Н— нейрон, возбуждаемый афферентными импульсами; 1 — нервные окончания, возбуждающие тормозной нейрон 7; 2— аксон, проводящий деполяризующие импульсы к телу нейрона Н, 3— аксоны тормозного нейрона, гиперполяризующие пресинаптические окончания возбуждающего нейрона
Постсинаптическое торможение.Наибольшее распространение в ЦНС имеет механизм постсинаптического торможения, которое осуществляется специальными тормозными вставочными нервными клетками — например, клетками Реншоу в спинном мозге или клетками Пуркинье (грушевидные нейроны) в коре мозжечка. Особенность тормозных клеток состоит в том, что в их синапсах
|
Реципрокное торможение.Механизм постсинаптического торможения лежит в основе таких видов торможения, как реципрокное и латеральное. Реципрокное торможение является одним из физиологических механизмов координации деятельности нервных центров. Попере
менно реципрокно тормозятся в продолговатом мозге центры вдоха и выдоха, прессорный и депрессорный сосудодвигательные центры и др. Особенно отчетливо реципрокное торможение проявляется на уровне спинного мозга при осуществлении строго координированных двигательных актов (ходьба, бег, чесание и др.). На уровне сегментов спинного мозга возбуждение группы мотонейронов, вызывающих сокращение мышц сгибателей, сопровождается реципрокным торможением другой группы мотонейронов, приводящих к расслаблению мышц разгибателей (рис. 3.13).
Существует два объяснения механизма спинального реципрокного торможения. Согласно первому объяснению, на пути от афферентного волокна к мотонейронам мышцы разгибателя при воз-
Вставочный тормозной нейрон
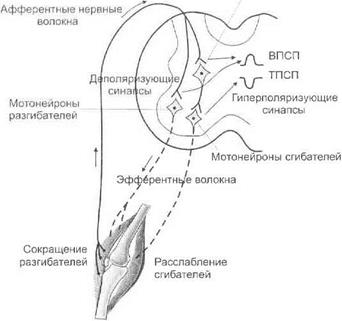
Рис. 3.13.Реципрокное торможение на уровне сегментов спинного мозга
буждении мотонейронов мышцы сгибателя включается механизм умножения импульсов. В результате мотонейроны разгибателей получают высокочастотную импульсацию, которая приводит их в состояние пессимума — торможения.
Согласно второй точке зрения, на пути к мотонейронам мышц разгибателя включаются вставочные тормозные нейроны, продуцирующие тормозной медиатор.
Латеральное торможение.При латеральном торможении активность нейронов или рецепторов, расположенных рядом с возбужденными нейронами либо рецепторами, прекращается. Механизм латерального торможения обеспечивает дискриминаторную способность анализаторов. Так, в слуховом анализаторе латеральное торможение обеспечивает различение частоты звуков, в зрительном анализаторе — резко увеличивает контрастность контуров воспринимаемого изображения, а в тактильном анализаторе — способствует дифференцировке двух точек прикосновения.
Латеральное торможение в значительной степени связано с механизмами возвратного торможения.
3.5. МЕТОДЫ ИЗУЧЕНИЯ ФУНКЦИЙ ЦНС
Метод хирургических экстирпаций.Включает удаление различных структур мозга, механическое их разрушение или коагуляцию анодом постоянного тока. Функциональное выключение структур мозга достигается их охлаждением или анодной поляризацией. Недостатком этого метода является кровоизлияние в зону разрушения и раздражение образующейся рубцовой тканью.
Разновидностью является метод локальной экстирпация ткани мозга через вживленные канюли.
Метод электрического и химического раздражения различных структур мозга.Включает раздражение обнаженных структур мозга. Впервые прямое раздражение коры головного мозга осуществили немецкие исследователи Г. Фрич и Е. Гитциг. Этот метод также грубый. Он сопровождается вскрытием мозговых оболочек, нарушением внутримозгового давления и др.
Широкое распространение получил стереотаксический метод введения раздражающих электродов в определенные точки мозга через трепанационные отверстия в черепе.
Стереотаксическая техника.Для введения электродов в мозг голова животных под наркозом закрепляется в стереотаксическом аппарате фиксаторами, введенными в ушные проходы, за нижние края глазниц или верхние челюсти.
Для ориентации электродов в мозге используют стереотаксические атласы для определенных видов животных. В атласах представлены серийные срезы мозга во фронтальной, горизонтальной и сагиттальной плоскостях.
Отсчет производится от нулевых плоскостей. У кошки, например, фронтальная нулевая плоскость проходит через наружные слуховые проходы. Горизонтальная нулевая плоскость — на 10 мм выше наружных слуховых проходов Нулевая сагиттальная плоскость располагается по сагиттальному шву.
Для введения электрода в мозг по атласу выбирается фронтальная плоскость, на которой необходимая структура выражена наиболее отчетливо (например, для вентромедиального гипоталамуса F + 12 мм, L = 2 мм, Н = 12 мм).
Погружаемый электрод фиксируется в электрододержателе. Прежде всего устанавливают держатель в таком положении, чтобы кончик электрода располагался над нулевой фронтальной плоскостью. Затем продвигают держатель
электрода на 12 мм вперед и на 2 мм в сторону. В этой точке в кости черепа сверлится отверстие, и электрод погружают в мозг на глубину 12 мм. Дополнительный контроль локализации кончиков электродов в мозге осуществляется с помощью рентгеноскопии
Для клинических целей существуют стереотаксисы для человека со специальными атласами.
Раздражение структур мозга может осуществляться изолированными, за исключением кончиков, биполярными электродами (расстояние между их кончиками 0,5 мм и меньше) или униполярно, когда индифферентный электрод располагается над носовой пазухой или в мышце.
Для химической стимуляции структур мозга используют каню- ли-хемотроды. Раздражение мозга может осуществляться контактно или путем телестимуляции.
Регистрация электрической активности головного мозга.Регистрация биопотенциалов с поверхности головы или черепа — электроэнцефалография.
Электроэнцефалограмма (ЭЭГ) отражает спонтанную активность мозговых структур: нейронов, синапсов, глии и межклеточного вещества. После смерти субъектов спонтанная ЭЭГ исчезает. Изменения ЭЭГ могут быть вызванными при нанесении раздражений на субъекта или непосредственно на мозговые структуры. Отведение ЭЭГ может быть униполярным и биполярным.
Электрическая активность, регистрируемая с поверхности мозга, — электрокортикограмма.
Типы ЭЭГ:
• Медленные, высокоамплитудные колебания a-волны, амплитудой 70—100 мкв, частотой 8—12 в секунду.
a-волны регистрируются в состоянии покоя, после еды, в начальных стадиях сна, при закрывании глаз.
• Частые, низкоамплитудные колебания P-волны, амплитудой 5—10 мкв, частотой 30—60 в секунду.
p-волны регистрируются при бодрствовании, при эмоциональном возбуждении, при любых формах афферентного воздействия. Реакция смены высокоамплитудных медленных ЭЭГ колебаний на низкоамплитудную, высокочастотную активность получила название реакция пробуждения, реакция блокады a-ритма или реакция десинхронизации (рис. 3.14).
▲
77 I о,1 мВ
Рис. 3.14.Реакция десинхронизации ЭЭГ у наркотизированной уретаном кошки при электрической стимуляции седалищного нерва. Момент нанесения раздражения отмечен стрелкой
• Упорядоченный 9-ритм, амплитудой 70-80 мкв, частотой 4—6 в секунду. Характерен для состояния эмоционального напряжения субъекта. Как правило, регистрируется в гиппокампе, перегородке мозга и в коре головного мозга.
• Медленный, высокоамплитудный 8-ритм, амплитудой 100- 150 мкв, частотой 0,5—4 в секунду. Проявляется в состоянии глубокого сна и при опухолевых заболеваниях головного мозга.
• Спайкообразная активность. Амплитуда колебаний до 200— 300 мкв. Частота спайков варьирует. Проявляется при эпилепсии. Кроме того, эпилептиформная активность возникает в лимбических структурах головного мозга, в первую очередь в гиппокампе, при воздействии на субъектов, например, электромагнитных полей (рис. 3.15).
Теории происхождения электрической активности головного мозга.Некоторые исследователи рассматривают высокоамплитудную активность головного мозга как результат синхронизации спайков отдельных нейронов, распространяющихся по круговым нейронным цепям (Э. Эдриан).
Другие авторы считают, что медленная ЭЭГ-активность— результат суммации локальных синаптических потенциалов. При этом полагают, что частота колебаний ЭЭГ-волн определяется синаптической задержкой: при увеличении числа нейронов, образующих нейрональные цепи, ЭЭГ замедляется (Дж. Экклс).
Существует точка зрения, что корковая ЭЭГ определяется циркуляцией возбуждений между корой и таламусом (X. X. Джаспер,
Бета-ритм
|
|

 |
|
|
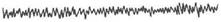
 |
|
|
 |
X. Т. Чанг и др.). Однако спонтанная ЭЭГ регистрируется и в изолированной коре, что делает более убедительными теории коркового происхождения ЭЭГ.
Все большее подтверждение находят электротонические теории происхождения электрической активности головного мозга. При этом полагают, что в апикальных аксодендритических эфаптических синапсах формируются суммирующиеся медленные потенциалы, образующие электрические поля (А. И. Ройтбак).
Обнаружено, что частота колебаний ot-ритма соответствует колебаниям электромагнитного поля Земли. Показано, что эпилептиформные разряды проходят через перерезанный спинной мозг (Ф. Бремер).
Метод функциональной дегенерации.При перерезке аксонов, например, пирамидных нейронов коры головного мозга, наблюдается дегенерация связанных с ними синапсов и нейронов нижележащих отделов мозга.
Метод пероксидазы хрена.Введение пероксидазы хрена в определенные структуры мозга вызывает реакцию в связанных с этими структурами других структурах мозга.
Стрихнинная нейронография.При накладывании стрихнина на определенные участки мозга спайковая активность регистрируется не только в пункте аппликации, но и в связанных с ним структурах мозга.
Аппликация химических веществ, микроионофорез.Аппликация блокаторов или активаторов синаптической передачи может осуществляться непосредственно на кору мозга или путем их подведения через хемотроды к различным глубинным структурам мозга. Вещества могут апплицироваться к отдельным нейронам мозга через микроканюли путем электрического отталкивания зарядов — микроионофорез.
Микродиализ.Осуществляется путем забора микродоз жидкостей из определенных структур мозга при различных состояниях экспериментальных животных с помощью специальных микропомп.
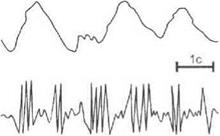
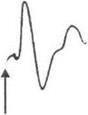
Метод вызванных потенциалов.При нанесении одиночных раздражений на периферические рецепторы или афферентные нервные стволы в проекционных зонах коры мозга и в различных подкорковых образованиях регистрируются вызванные потенциалы(рис. 3.16).
Вызванные потенциалы в проекционных зонах коры головного мозга имеют следующие фазы: первичные положительный и отрицательный потенциалы и вторичные потенциалы. Возникновение первичного положительного потенциала связывают с возбуждением аксосоматических синапсов на нейронах 3—4-го слоя коры, а первичного отрицательного потенциала — с возбуждением аксоден- дритных синапсов.
 | |||||||
 | |||||||
 | |||||||
 | |||||||
Рис. 3.16.Вызванный потенциал в зрительной коре головного мозга
в ответ на одиночную вспышку света
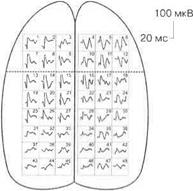
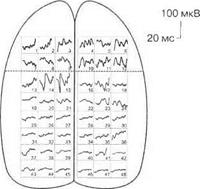
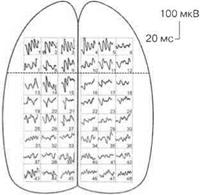
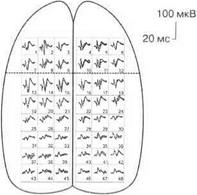
При различных видах наркоза выявляются различные карты распределения, например, соматосенсорных вызванных потенциалов, по коре головного мозга, что указывает на избирательное действие различных наркотических веществ на синаптические образования мозга (рис. 3.17).
Вызванные потенциалы, как правило, блокируются на фоне реакции десинхронизации ЭЭГ (рис. 3.18).
Микроэлектродные исследования.Для микроэлекгродных исследований электрической активности отдельных нейронов мозга применяют металлические или стеклянные микроэлектроды, заполненные раствором хлористого натрия. Диаметр кончиков микроэлектродов — 1—0,5 мкм.
В микроэлекгродных исследованиях используют методы внеклеточного и внутриклеточного отведения биопотенциалов.
Магнитно-резонансная томография.При облучении головного мозга электромагнитным полем диполи жидкостей мозга (например, молекулы воды) принимают направление облучающего поля. При выключении внешнего магнитного поля диполи возвращаются в исходное положение, при этом возникает магнитный сигнал, который улавливается специальными датчиками и регистрируется в виде графика на компьютере. Поскольку внешнее магнитное поле можно сделать плоским, то таким образом можно «резать» головной мозг послойно. Метод позволяет обнаруживать в головном мозге опухолевые образования и зоны нарушения мозгового кровообращения.
Позитронно-эмиссионная томография.Исследование основано на введении в мозговой кровоток позитрон-излучающих корот-
 |
|
 |
|
 |
|
 |
| ||||
| ||||
коживущих изотопов. Данные о распределении радиоактивности в мозге калькулируются за определенные отрезки времени на компьютере и затем реконструируются в трехмерный образ.
Метод позволяет наблюдать в различных отделах мозга очаги возбуждения при умственной деятельности испытуемых.
Моделирование функций мозга. Впоследние годы широкое распространение получило компьютерное моделирование функций
СМК
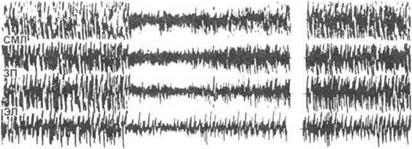

Рис. 3.18.Блокада зрительного вызванного потенциала на фоне ноцицептивного раздражения
мозга. Построены модели нервных сетей, выполняющие отдельные функции мозга. Создан «детектор интеллекта» (Е. А. Умрюхин), позволяющий определять индивидуальные свойства различных стадий системной организации психической деятельности человека.
3.6. ФИЗИОЛОГИЯ СПИННОГО МОЗГА
3.6.1. Роль спинного мозга в целостной деятельности организма
Спинной мозг — основной канал проведения афферентной (кожной, температурной, проприоцептивной и болевой) сигнализации от периферических рецепторов к высшим отделам ЦНС. При нарушении проведения афферентных импульсов по спинному мозгу (например, при спинной сухотке) человек при закрытых глазах не может совершать движения (атаксия). Мать, например, не может удерживать на руках ребенка. Спинной мозг — основной канал обратной афферентации о результатах двигательных актов.
Спинной мозг — также ведущий канал проведения возбуждений из ЦНС к мотонейронам и вставочным нейронам боковых рогов, формирующих двигательные и вегетативные реакции (см. разд. 3.17).
Проводящие пути спинного мозга.Через спинной мозг пролегают основные афферентные пути от периферических кожных, проприоцептивных и висцеральных рецепторов в высшие отделы ЦНСи эфферентные пути распространения возбуждений из ЦНСк мышцам и висцеральным нервам.
Ведущие афферентные пути:
• путь Голля и Бурдаха;
• спинно-таламический тракт;
• спинно-мозжечковый тракт.
Эфферентные пути:
• пирамидный тракт;
• экстрапирамидные пути:
— ретикулоспинальный тракт;
— руброспинальный тракт;
— тектоспинальный тракт;
— вестибулоспинальный тракт.
Пирамидный тракт имеет обширные коллатерали к различным отделам ЦНС.
Проводящие пути спинного мозга подробно описываются в курсе нормальной анатомии.
Спинальный шок.Наблюдается после перерезки спинного мозга. При этом все функции спинного мозга исчезают. У лягушек рефлекторные реакции спинного мозга быстро (через 10—15 мин) восстанавливаются. У человека восстановления функций разорванного спинного мозга практически не происходит.
Теории спинального шока. Ф. Гольтц рассматривал спинальный шок как результат раздражения. Тем не менее спинальный шок также наблюдается при холодовой блокаде спинного мозга (Ч. Шеррингтон, Г. Тренделенбург). Повторная перерезка спинного мозга ниже первой перерезки также не вызывает спинального шока. Все это указывает на то, что спинальный шок возникает вследствие отделения спинного мозга от вышележащих отделов головного мозга.
Микроэлектродные исследования показали, что при спинальном шоке мотонейроны не страдают. При этом повреждаются вставочные нейроны, вследствие чего отсутствуют реакции на афферентные воздействия.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|