- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Вегетативная нервная система 3 страница
3.6.2. Собственные функции спинного мозга
• Спинальные рефлексы.
• Координация двигательной деятельности организма, в частности функций спинальных мотонейронов.
• Вегетативные реакции.
• Мочевыделение и дефекация.
Животные с перерезанным спинным мозгом на границе с продолговатым получили название спинальных животных.
У спинальных животных:
• отсутствует самостоятельное дыхание;
• низкое кровяное давление, сниженный сосудистый тонус;
• утрачивается способность поддерживать постоянство температуры тела;
• исчезают все формы сложной целенаправленной деятельности (пищевая, половая, оборонительная);
• при перерезках ниже люмбальных отделов наблюдается паралич анальных и мочеиспускательных сфинктеров.
Вегетативные функции спинного мозга.На уровне спинного мозга осуществляются вегетативные функции:
• дефекация;
• мочеиспускание;
• регуляция сосудистого тонуса;
• эрекция и эякуляция;
• потоотделение.
3.6.3. Спинальные механизмы координации двигательной деятельности
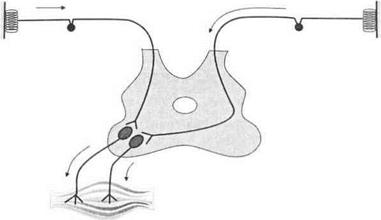
Моно- и полисинаптические реакции спинного мозга.Моноси- наптически афферентные пути непосредственно переключаются на мотонейроны. Примером моносинаптической реакции является коленный рефлекс. Полисинаптические реакции включают несколько вставочных нейронов. Они могут быть моно- и полисегментарными (рис. 3.19).
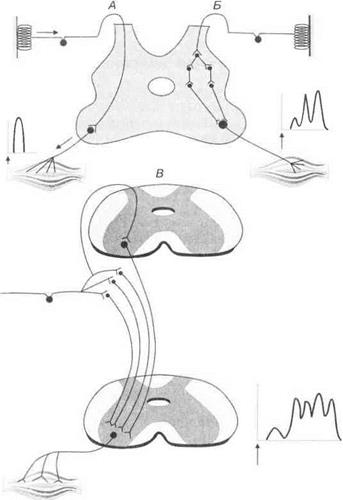
Рис. 3.19.Моносинаптическая (А), полисинаптическая (5) и полисег- ментарная (В) рефлекторные дуги
Моторное поле спинальных двигательных нейронов.Каждый мотонейрон иннервирует определенное число мышечных волокон (моторные единицы). Совокупность нейронов, иннервирующих целую мышцу, составляет моторное поле мыищы (рис. 3.20).
 |
|
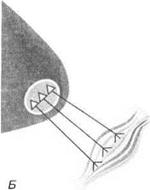 |
Фракционирование моторного поля.Каждое афферентное волокно обслуживает только часть моторного поля (рис. 3.21). На раздражение чувствительного нерва мышца поднимает меньший груз, чем на раздражение двигательного нерва. Каждый чувствительный нерв возбуждает только часть моторного поля мышцы.
 |
|
Окклюзия (закупорка).Вытекает из принципа фракционирования моторного поля. При электрическом раздражении одного чувствительною нерва мышца поднимает, например, 600 г груза, при раздражении другого чувствительного нерва — 400 г. При одновременном раздражении обоих чувствительных нервов мышца поднимает груз только весом 800 г.
Это объясняется тем, что часть афферентных волокон конвергирует на одни и те же мотонейроны спинного мозга (рис. 3.22). Чем больше конвергенция сенсорных возбуждений, тем более выражена окклюзия.
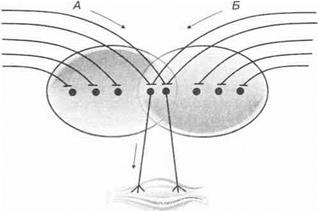
Рис. 3.22.В основе феномена окклюзии лежит конвергенция афферентных возбуждений (Д и Б) на одни и те же мотонейроны спинного мозга
Пространственная суммация спинальных возбуждений.Феномен окклюзии обусловливается в спинном мозге пространственной суммацией возбуждений. Она проявляется при подпороговых раздражениях чувствительных нервов, когда раздражение каждого нерва по отдельности не вызывает мышечного сокращения, но их совместное раздражение приводит к сокращению мышц.
Реципрокность эффекторных возбуждений.Главная роль в координации двигательной деятельности спинного мозга принадлежит конечным мотонейронам.
При раздражении чувствительных нервов в одних случаях происходит сокращение сгибательных и одновременное расслабление разгибательных мышц, и наоборот.
Как указывалось выше, существует два объяснения феномена реципрокности. Считают, что торможение экстензорных нейронов при возбуждении флексорных происходит за счет включения вставочных нейронов и пессимума их мотонейронов (Г. Гассер). Другая точка зрения заключается в том, что во вставочных нейронах на пути к экстензорным мотонейронам выделяются тормозные медиаторы (глицин, [3-аланин и др.) (см. разд. 3.4.2).
Свойства конечных мотонейронов.Конвергенция множества афферентных и эфферентных возбуждений с периферии и из высших отделов ЦНС на мотонейронах (рис. 3.23). Благодаря этому одни и те же мотонейроны могут участвовать во многих реакциях.
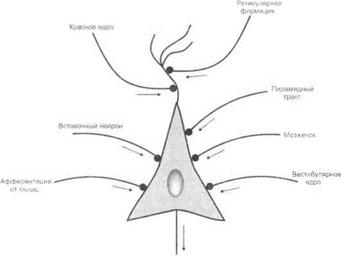
Рис. 3.23.Множественная конвергенция афферентных возбуждений на спинальном мотонейроне
Ч. Шеррингтон сформулировал «принцип воронки», или «борьбы за конечный путь». Однако по существу никакой борьбы на спинальных мотонейронах нет. Их эффекторная импульсация определяется суммарной разностью потенциалов между «светлым пятнышком» и суммой дендритных потенциалов, а также, возможно, спецификой постсинаптических химических реакций.
Деятельность конечных мотонейронов спинного мозга контролируется также антидромно за счет возвратных коллатералей их аксонов и тормозных клеток Реншоу, выделяющих тормозные медиаторы.
Синдром Броун—Секара.Проявляется при половинной перерезке спинного мозга. При этом на стороне перерезки отмечается:
• вялый паралич (не устраняются неперекрещивающиеся волокна пирамидного тракта);
• исчезает проприоцептивная и тактильная чувствительность в результате перерезки путей Голля и Бурдаха.
На противоположной стороне отмечается потеря болевой и температурной чувствительности.
3.7. ФИЗИОЛОГИЯ ПРОДОЛГОВАТОГО МОЗГА И ВАРОЛИЕВА МОСТА
Животные с перерезками головного мозга по переднему краю варолиева моста называются бульбарными.
Бульбарные животные самостоятельно дышат, поддерживают постоянный уровень артериального давления. Однако они не могут поддерживать постоянство температуры тела и передвигаться.
Бульбарные реакции.Проявляются при воздействиях на бульбарных животных внешних раздражителей. Это пищевые и защитные реакции.
Пищевые реакции: глотание, сосание, жевание, рефлекторная деятельность пищеварительного тракта (слюноотделение, секреция желудка и других пищеварительных органов).
Защитные реакции: чихание, кашель, рвота, мигание (моргание), слезоотделение.
Указанные реакции сложно координированы. За счет объединения нескольких ядер нервов они охватывают комплекс исполнительных мышечных и вегетативных реакций.
Каждая из этих реакций имеет специфическое рецепторное (рефлексогенное) поле. Например, глотание формируется при пищевом тактильном раздражении задней стенки ротовой полости, мигание — при раздражении конъюнктивы глаза и т.п.
Каждую бульбарную реакцию вызывает специфический раздражитель. Так, механическое раздражение глотки вызывает рвоту, раздражение той же области пищей — глотание.
Автоматическая деятельность бульбарных нервных центров.Нейроны продолговатого мозга определяют автоматию вдоха-выдоха.
Причиной автоматии дыхательных бульбарных центров являются изменение в них внутренних метаболических процессов, корригирующее влияние блуждающих нервов и влияние двуокиси углерода (см. разд. 5.3).
Тонические бульбарные центры.Благодаря афферентным нервным и гуморальным воздействиям бульбарные центры оказывают постоянные нисходящие влияния на тонус артериол, в результате чего поддерживается оптимальный для метаболизма тканей уровень кровяного давления (см. разд. 5.2.12).
Посредством эфферентных волокон блуждающие нервы бульбарных центров оказывают влияние на тоническое напряжение мышц желудка и стенок кишечника при голоде.
Сахарный укол.При локальном уколе в дно 4-го желудочка К. Бернар обнаружил резкое увеличение содержания глюкозы в крови.
Регуляция деятельности спинного мозга.При электрическом раздражении вентромедиальных отделов ретикулярной формации ствола мозга наблюдается торможение:
• коленного рефлекса;
• кортикально вызванных движений;
• передачи возбуждения по у-афферентным волокнам.
При этом утрачивается общий мышечный тонус — животные распластываются.
При раздражении дорсолатеральных участков ретикулярной формации ствола мозга облегчаются коленный рефлекс и кортикально вызванные движения.
Увеличивается мышечный тонус мышц сгибателей и разгибателей (X. Мэгун) (рис. 3.24).
Указанные нисходящие влияния осуществляются через вестибулоспинальный тракт.
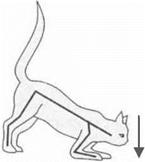
Децеребрационная ригидность.Перерезка на уровне переднего края варолиева моста вызывает у животных усиление тонуса мышц
|
 |
Рис. 3.24.Облегчающие и тормозные нисходящие влияния ретикулярной формации продолговатого и среднего мозга на спинальные реакции
Рис. 3.25.Децеребрационная ригидность
разгибателей (рис. 3.25). Животные не сохраняют равновесия, падают. Это явление получило название децеребрационная ригидность. Полагают, что децеребрационная ригидность связана с устранением нисходящих влияний мозжечка и красных ядер среднего мозга на бульбарные центры.
3.8. ФИЗИОЛОГИЯ СРЕДНЕГО МОЗГА
Животные с перерезкой мозга на уровне передних бугорков четверохолмия получили название мезенцефальных животных.
Мезенцефальные животные отличаются от спинальных регуляцией мышечного тонуса и положения тела в пространстве. У мезенцефальных животных отсутствует явление децеребрационной ригидности. Красные ядра, локализованные в среднем мозге, оказывают тормозное влияние на вестибулярные ядра Дейтерса, к которым распространяется афферентация от ушных лабиринтов и мышечных проприоцепторов по вестибулоспинальному тракту (рис. 3.26). Разрушение красных ядер вызывает у животных выраженную децеребрационную ригидность. Раздражение красных ядер приводит к резкому понижению мышечного тонуса — животные провисают «как мешок».
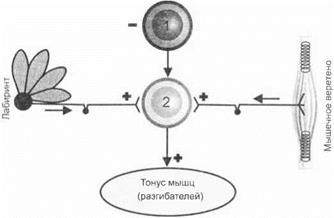
Рис. 3.26.Схема взаимоотношения красного ядра (1) и вестибулярного ядра Дейтерса (2) в регуляции мышечного тонуса (децеребрационной ригидности)
Децеребрационная ригидность определяется также афферента- цией, поступающей к ядрам Дейтерса от проприоцепторов мышц и вестибулярного аппарата. Перерезка задних корешков спинного мозга и вестибулярных нервов также устраняет децеребрационную ригидность.
При отсутствии тормозящих влияний красных ядер на уровне заднего мозга происходит такая «раскладка» центральных возбуждений, при которой избирательно повышается активность мышц разгибателей.
3.8.1. Собственные функции среднего мозга
Значительный вклад в исследование функций среднего мозга внесен голландским физиологом Р. Магнусом.
Регуляция позы(позные рефлексы) осуществляется тем лучше, чем ниже животные в эволюционном ряду. Мезенцефальные кролики и лягушки сидят и прыгают как обычно.
Позные рефлексы (они же рефлексы положения, познотониче- ские рефлексы) включают:
1. Статические рефлексы (миостатика).Определяют поддержание определенной позы в пространстве при лежании, сидении и стоянии животных за счет изменения при этом мышечного тонуса.
Статические рефлексы связаны с:
• Раздражением лабиринтов вестибулярного аппарата.
Р. Магнус и Де-Клейн загипсовывали животных так, что их голова не могла смещаться по отношению к туловищу. Вращение таких животных вокруг их продольной и поперечной оси вызывало выраженные изменения тонуса мышц сгибателей или разгибателей. При вращении вправо, например, вытягивались левые лапы и поджимались правые.
• Положением головы по отношению к туловищу.
При поднятой голове усиливается тонус разгибателей передних и сгибателей задних конечностей, а при опущенной голове — тонус мышц разгибателей задних конечностей и сгибателей передних конечностей (рис. 3.27).
При поворотах головы в сторону повышается тонус разгибателей передней конечности на стороне, куда повернута голова, и мышц сгибателей передней конечности противоположной стороны.
• Сигнализацией от рецепторов кожи, мышц и суставов всего тела.
• Зрительной афферентацией.
2. Установочные (выпрямительные) рефлексы.Определяют возвращение животного из неустойчивого положения в устойчивое, а также смену поз.
 | |||
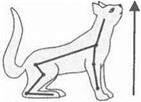 | |||
|
Эти реакции также определяются:
• раздражением лабиринтов вестибулярного аппарата;
• афферентацией от шейных мышц;
• афферентацией от кожных рецепторов всей поверхности тела.
Основной фактор при смене позы — изменение положения головы. Лежащее животное первым делом поднимает голову и устанавливает ее в горизонтальном положении. Вслед за этим немедленно за счет шейно-тонических рефлексов и афферентации от рецепторов мышц и кожи перераспределяется мышечный тонус, определяющий вставание животного. Если на мезенцефальное животное надавить сверху доской, то голова его немедленно падает.
3. Изменение тонуса мышц в связи с движениями животных (статокинетические рефлексы).При движении животных тонус мышц все время перераспределяется. Это отчетливо проявляется при помещении собаки на платформу, разделенную на четыре площадки, на каждой из которых помещается одна лапа животного. При поднятии животным одной лапы изменяется мышечный тонус остальных трех лап. Явление получило название позиционного возбуждения(М.Ф. Корякин) (см. также разд. 7.2).
3.8.2. Функции четверохолмий
Нейроны ядер четверохолмий определяют такие компоненты ориентировочных реакций, как:
• прислушивание;
• перемещение глазных яблок;
• регуляцию просвета зрачка;
• поворот ушных раковин у животных;
• сторожевые реакции при внезапных световых или звуковых раздражениях;
• старт-реакции — немедленную мобилизацию мышечного аппарата к действию.
Верхние бугры четверохолмия — первичный зрительный подкорковый центр, обеспечивающий поворот головы и глаз в сторону светового раздражителя, фиксацию взора и слежение за движущимися объектами. Нижние бугры четверохолмия — первичный слуховой подкорковый центр, обеспечивающий ориентировочный слуховой рефлекс.
Корковые влияния могут усиливать или ослаблять эти реакции (см. также разд. 6.3.1. и 6.3.2).
3.8.3. Черная субстанция
Расположенные здесь нейроны симпатической нервной системы иннервируют красные мышечные волокна. При поражениях черной субстанции нарушается пластический тонус поперечно-полосатых мышц. Больные могут часами лежать с поднятой головой.
Черная субстанция участвует в регуляции актов жевания, глотания и их последовательности.
В черной субстанции сосредоточены дофаминовые нейроны, участвующие в организации эмоционального поведения и в регуляции тонких движений (например, пальцев рук).
3.8.4. Серое вещество вокруг сильвиева водопровода
Нейроны серого вещества обладают рецепторной функцией по отношению к спинномозговой жидкости. Участвуют в поддержании внутренней среды на постоянном уровне.
Проводниковая функция среднего мозга.Осуществляется через ножки мозга. Особые функции — у ретикулярной формации среднего мозга (см. разд. 3.9).
3.9. ФИЗИОЛОГИЯ РЕТИКУЛЯРНОЙ ФОРМАЦИИ СТВОЛА МОЗГА
Ретикулярная (сетчатая) формация охватывает значительное пространство ствола мозга. Она включает: латеральные ретикулярные ядра, ретикулярные вентральные ядра, ядра покрышки мозга (ядра Бехтерева), парамедиальные ретикулярные ядра, ретикулярные гигантоклеточные ядра, оральные ретикулярные ядра моста, каудальные ретикулярные ядра моста; ретикулярные мелкоклеточные ядра.
Некоторые исследователи включают в состав ретикулярной формации ствола мозга задние отделы гипоталамуса и вентральные ядра таламуса.
Ретикулярная формация морфофункционально тесно связана:
• с мозжечком;
• со спинным мозгом;
• с вышележащими отделами головного мозга.
Функции ретикулярной формации:
1. Раздражение ретикулярной формации через вживленные электроды пробуждает спящее животное.
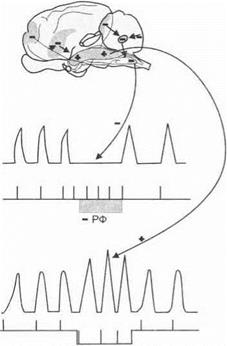
2. При электрическом раздражении ретикулярной формации у животных, находящихся под наркозом, наблюдается реакция активации ЭЭГ (рис. 3.28).
А f л 101м3
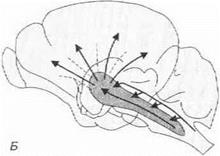 Рис. 3.28.Изменение электрической активности коры головного мозга у животного под уретановым наркозом при раздражении ретикулярной формации ствола мозга (А). Ретикулярная формация оказывает на кору головного мозга генерализованные неспецифические восходящие активирующие влияния (Б)
Рис. 3.28.Изменение электрической активности коры головного мозга у животного под уретановым наркозом при раздражении ретикулярной формации ствола мозга (А). Ретикулярная формация оказывает на кору головного мозга генерализованные неспецифические восходящие активирующие влияния (Б)
3. При двустороннем разрушении структур ретикулярной формации ствола головного мозга у животных наблюдается беспробудный сон (Д. Линдсли).
Таким образом, ретикулярная формация ствола мозга оказывает на кору больших полушарий восходящие активирующие влияния. Эти влияния генерализованные. Они адресуются практически ко всем отделам головного мозга.
Восходящие активирующие влияния ретикулярной формации.Выявление генерализованных восходящих активирующих влияний ретикулярной формации ствола мозга заставило изменить классические представления о поступлении информации в кору головного мозга. Классические представления о проводниковом распространении афферентных возбуждений по лемнисковым путям дополнились генерализованными влияниями на кору головного мозга.
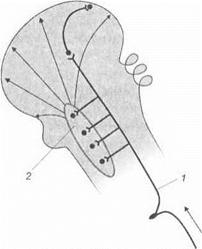
|
Тонические активирующие влияния ретикулярной формации формируются за счет постоянного поступления к ее структурам афферентных нервных импульсов от различных органов чувств и периферических рецепторов, а также благодаря наличию в ней многочисленных аппаратов мультипликации импульсов, «нейронных ловушек» и циклических (реверберирующих) взаимодействий между различными ее отделами и другими структурами мозга.
Кроме того, активирующее действие ретикулярной формации ствола мозга определяют гуморальные факторы. Показано, что инъекции адреналина непосредственно в ростральные отделы ретикулярной формации вызывают активацию ЭЭГ (М. Фогт). Такое же действие оказывает аппликация СО, к структурам ретикулярной формации среднего мозга (Е. Л. Голубева).
Таким образом, любые сенсорные и гуморальные факторы активируют ретикулярную формацию ствола головного мозга. Это указывает на неспецифические восходящие активирующие влияния ретикулярной формации на кору головного мозга и его другие образования.
Наряду с этим установлено (В. Г. Агафонов), что ретикулярная формация избирательно чувствительна к различным наркотическим веществам.
Так, оказалось, что под уретановым наркозом при раздражении седалищного нерва отчетливо проявляются восходящие активирующие влияния ретикулярной формации на кору головного мозга. В то же время они не проявляются у животных, находящихся под нембуталовым наркозом. Все это указывает на химическую неоднородность структур ретикулярной формации ствола мозга.
Подтверждением этого являются эксперименты, показавшие, что неблокируемые уретаном восходящие активирующие влияния ретикулярной формации на кору головного мозга при ноцицептивных раздражениях животных блокировались дополнительным введением им а-адреноблокатора — аминазина. Все это также указывает на наличие в составе ретикулярной формации структур, определяющих состояние бодрствования, и химически отличных структур, определяющих состояние боли.
Показано, что аминазин избирательно устраняет активацию условно-рефлекторного оборонительного состояния у животных и не влияет на их пищевые условные рефлексы (А. И. Шумилина).
Все это позволило сформулировать представления о специфических восходящих активирующих влияниях ретикулярной формации на кору головного мозга при реакциях различного биологического качества (П. К. Анохин).
Специальные эксперименты продемонстрировали, что специфика восходящих активирующих влияний ретикулярной формации на кору головного мозга при реакциях различного биологического качества определяется дополнительными влияниями структур
гипоталамуса и лимбических образований головного мозга (см. разд. 8.5).
Все это заставляет говорить о восходящих активирующих влияниях подкорковых образований на кору больших полушарий.
В структурах ретикулярной формации обнаружены адренергические, дофаминергические, холинергические нейроны, что еще раз подтверждает представления о химической гетерогенности структур ретикулярной формации ствола головного мозга.
Как указывалось выше, структуры ретикулярной формации ствола мозга оказывают облегчающее и тормозное влияния на рефлекторные реакции спинного мозга.
Таким образом, ретикулярная формация — структура мозга, обеспечивающая тоническими влияниями другие отделы мозга, бодрствующее состояние субъектов, определяющая активные связи организма с внешней средой. Некоторые авторы (У. Пенфилд) даже считают, что ретикулярная формация ствола мозга определяет сознание человека (центрэниефалическая теория).
Открытие свойств ретикулярной формации ствола мозга изменило традиционные представления об организации корково-подкорковых соотношений, механизмах действия наркотиков и психофармакологических средств, механизмах боли и обезболивания, бодрствования и сна, мотиваций и эмоций, условных рефлексов и анализаторов.
3.10. ФИЗИОЛОГИЯ МОЗЖЕЧКА
Функции мозжечка связаны, главным образом, с организацией двигательных актов и регуляцией вегетативных функций.
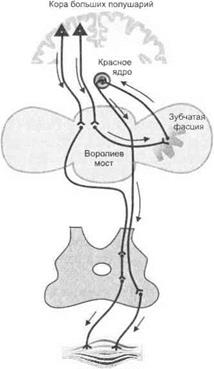
При осуществлении двигательного акта перемещающиеся части тела испытывают влияние инерционных сил, что нарушает плавность и точность выполняемого движения. Структуры мозжечка осуществляют коррекцию движения. В промежуточную часть мозжечка по коллатералям кортикоспинального тракта из лобных отделов коры головного мозга поступает информация о планируемом движении, а также соматосенсорная афферентация. В результате формируются потоки возбуждения к красному ядру и стволовым двигательным центрам, обеспечивающие взаимную координацию позных и тонических движений, а также коррекцию выполняемого
движения. Мозжечок также получает импульсы из коры больших полушарий через фронто-понто-церебеллярный тракт. Из зубчатого ядра мозжечка некоторые волокна посредством зубчато-рубро- таламо-кортикального тракта проецируются в таламус и оттуда в фронтальную кору через хвостатое ядро.
Мозжечок, таким образом, контролирует функцию фронтальной коры посредством механизма обратных связей.
Мозжечок определяет построение быстрых баллистических целенаправленных движений (например, бросание мяча в цель, прыжок через препятствие, игра на фортепиано). В таких случаях коррекция по ходу выполнения движения невозможна из-за малых временных параметров, и баллистическое движение будет выполнено только по заранее заготовленной программе. Программа формируется в коре больших полушарий мозга. Отсюда возбуждения поступают к зубчатому ядру мозжечка. Импульсации мозжечка контролируют выполнение движений. В течение всей жизни человека в мозжечке непрерывно формируются двигательные программы с сохранением информации, позволяющей пирамидной и экстрапирамидной системам формировать необходимый комплекс двигательных импульсов, под действием которых выполняется необходимое движение.
Функции мозжечка подробно изучены на безмозжечковых собаках (Л. Лючиани). При удалении мозжечка у собак наблюдается мышечный гипертонус, похожий на децеребрационную ригидность, за счет ослабления влияний красных ядер среднего мозга. В дальнейшем развиваются следующие симптомы:
• Атония (гипотония) — снижение мышечного тонуса. Голова животных повисает на грудь.
• Абазия — потеря площади опоры. При этом ходьба осуществляется широко расставленными ногами (как при качке на палубе).
• Астазия — нарушение стояния. Усиленное качание из стороны в сторону. У людей астазия проявляется при нарушении функций мозжечка, при закрытых глазах и вытянутых вперед руках (проба Ромберга).
• Атаксия — нарушение движения. Походка становится расхлябанной, как у пьяного.
• Астения — повышенная утомляемость. При этом зубчатый тетанус выявляется при тех частотах раздражений мышц, при
которых обычно проявляется гладкий тетанус — повышается функциональная лабильность мышц.
• Наблюдается дрожание — статический и динамический тремор.
• Возрастает термопродукция.
В целом поражение мозжечка вызывает нарушение равновесия в покое и при движении. При этом субъекты теряют способность оценивать вес — на стороне поражения мозжечка вес оценивается как более легкий.
|
Мозжечок имеет только один эффекторный выход — через аксоны грушевидных клеток Пуркинье. Аксоны клеток Пуркинье заканчиваются преимущественно на нейронах крупноклеточной части латерального вестибулярного ядра (ядро Дейтерса). Через вестибулоспинальные пути осуществляется фазный и тонический контроль экстен- зорной (антигравитационной) мускулатуры.
Клетки Пуркинье оказывают на сегментарный аппарат спинного мозга тормозное влияние.
Иногда последующие влияния мозжечка тормозятся корковыми влияниями, тогда вся сила вкладывается в движение (например, при ударе кулаком).
Устранение мозжечковых влияний приводит к:
• размашистости движений (вместо круга больные рисуют изломанную форму);
• отсутствию точных движений;
• больные с поражением мозжечка не могут при закрытых глазах дотронуться до кончика носа (пальценосовая проба);
• изменению речи (сканирование, разрывание речи на необычных слогах);
• при откидывании головы назад субъект падает.
Наряду с регуляцией мышечного тонуса и равновесия мозжечок при взаимодействии с лимбическими структурами головного мозга участвует в регуляции вегетативных функций — дыхания, кровообращения, перистальтики желудочно-кишечного тракта, потоотделения.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|