- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Веин А.М., Наменецкая Б.И. – Память человека 2 страница
|
|

Проведенный анализ свидетельствует о том, что восприятие информации является сложным процессом, включающим активный отбор на основе сличения воспринятого с прошлым опытом и его эмоциональной оценкой, внимание, классификацию воспринятой информации, что подготавливает след к запоминанию.
В настоящее время накопились клинические наблюдения и экспериментальные факты, которые убедительно свидетельствуют о том, что вслед за восприятием информации идет период ее закрепления в памяти, фаза консолидации, без которой поступающая информация не может быть сформирована в стабильный след памяти. Эти данные, несомненно, свидетельствуют о том, что в осуществлении процесса консолидации принимают участие глубинные структуры мозга, и прежде всего образования гиппокампова круга, называемого также кругом Пейпеца. Он включает в себя, помимо образований гиппокампа, передние ядра таламуса и мамиллярные тела. Так, оказалось, что при двустороннем поражении структур гиппокампова круга у людей (см. главу VIII), а также при искусственном их разрушении у различных видов живот
ных выявляются резко выраженные своеобразные нарушения памяти, для которых характерна невозможность усвоения любого нового материала. Эти наблюдения позволили предположить особую роль этих образований мозга в процессах памяти. Почти 100-летний клинический опыт, а также многие экспериментальные факты свидетельствовали о появлении наиболее грубых, тотальных нарушений памяти только при поражении структур гиппокампова круга, относящихся к образованиям лимбической системы. Прежде чем перейти к более подробному изложению имеющихся наблюдений, целесообразно коротко остановиться на понятии лимбической системы мозга, се структурных и функциональных особенностях.
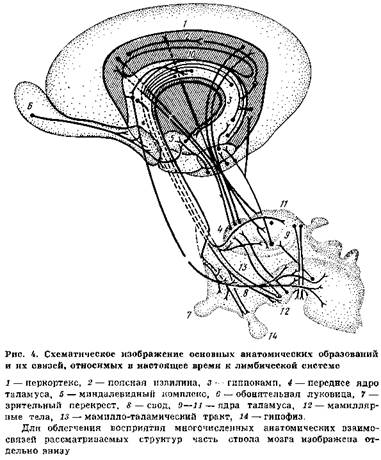
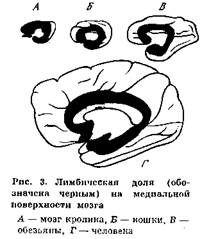 Лимбическая система является функциональным понятием, объединяющим структуры, которые принимают участие в организации сложных поведенческих актов. Эта система включает глубинные образования мозга, имеющие тесные функциональные связи. Центральные звенья лимбической системы — миндалевидный комплекс и гиппокамп, расположенные в глубине височной доли мозга. Гиппокамп тесно связан с другими анатомическими структурами (рис. 3), входящими в лимбическую систему, и образует вместе с ними упомянутый выше гиппокампов круг: гиппокамп — свод — перегородка — мамиллярные тела — передние ядра таламуса — поясная извилина — гиппокамп (рис. 4).
Лимбическая система является функциональным понятием, объединяющим структуры, которые принимают участие в организации сложных поведенческих актов. Эта система включает глубинные образования мозга, имеющие тесные функциональные связи. Центральные звенья лимбической системы — миндалевидный комплекс и гиппокамп, расположенные в глубине височной доли мозга. Гиппокамп тесно связан с другими анатомическими структурами (рис. 3), входящими в лимбическую систему, и образует вместе с ними упомянутый выше гиппокампов круг: гиппокамп — свод — перегородка — мамиллярные тела — передние ядра таламуса — поясная извилина — гиппокамп (рис. 4).
Лимбическая система регулирует функциональное состояние внутренних органов и желез внутренней секреции; поведенческие акты, связанпые с пищевой, половой функцией и эмоциональной сферами; принимает участие в регуляции сна и бодрствования, внимания, процессов памяти. Лимбическая система играет активирующую и модулирующую роль по отношению ко всем функциям
|
|
и таким образом осуществляет сложную интегративную деятельность мозга.
Функции в лимбической системе представлены глобально, топографически плохо дифференцируются, однако определенные отделы имеют специфические задачи в организации целостных поведенческих актов. Изучением функциональной роли этих образований мозга занимаются такие крупные нейрофизиологи, как П. К. Анохин, Мак Лин, Дельгадо, Прибрам, Граштьян и др.
В настоящее время можно считать доказанным, что гиппокамп как важнейшая часть лимбической системы играет существенную роль в регистрации информации. Однако общая функция лимбической системы осуществляется путем взаимодействия отдельных структур гиппо- кампова круга, связанных между собой прямыми путями (круг Пейпеца). Различие структурной организации этих образований позволило предположить и их различную функциональную роль в процессе регистрации информации, поскольку функция мозга тесно связана с его структурной организацией и, как выразился известный американский исследователь памяти Р. Сперри, «функция кристаллизуется в его структуре». Это предположение подтверждено рядом исследователей, в том числе О. С. Виноградовой с сотрудниками при регистрации нейронной активности отдельных образований лимбической системы. Проведенные исследования позволили считать, что лимбическая система может быть разделена по функциональному назначению на два класса структур: 1) обеспечивающих необходимую фоновую активность и отбирающих новую нужную информацию и 2) кодирующих воспринятую информацию.
Первая группа структур, к которым относится гиппокамп и связанные с ним определенные отделы ретикулярной формации, получает большой приток информации от ретикулярной формации и коры головного мозга. Эти структуры реагируют однотипно на разные стимулы и имеют большой процент нейронов, осуществляющих торможение определенных форм деятельности. Истинным раздражителем этих структур является степень новизны раздражителя. При постоянстве окружающей среды нейроны гиппокампа находятся в активном состоянии. При любых изменениях в окружающей обстановке разряд в этих структурах полностью гасится. При постоянстве внешней обстановки гиппокамп тормозит активность ретикулярной формации. Изменение обстановки приводит к растормаживанию ретикулярной формации, что обеспечивает мобилизацию внимания и регистрацию нового раздражителя. При повторном воздействии раздражителя растормаживапие ретикулярной формации происходит на все более короткое время и постепенно затухает. Одновременно с этим прекращается - и процесс регистрации. Таким образом осуществляется отбор информации для ее регистрации и поддерживается необходимый рабочий уровень всей системы.
Другие структуры лимбической системы — мамилляр- ные тела, передние ядра таламуса и часть лимбической коры могут быть отнесены ко второму классу структур, осуществляющих кодирование информации. Так, часть нейронов в мамиллярных телах, к которым приходят импульсы из гиппокампа, отвечают на раздражение уже не однотипно, а соответственно их длительности, проявляя способность к оценке временных параметров поступающей информации. Нейроны передних таламических ядер, к которым попадает импульсация из мамиллярных тел, работают уже с высокой избирательностью, так как активно отвечают на одни раздражители, хуже на другие и совсем не отвечают на третьи. Эти нейроны мономо- дальны и проявляют избирательность даже в пределах раздражителей одной модальности. Некоторые из этих нейронов слабо отвечают, например, на чистые тоны, но сильно и более устойчиво реагируют на «натуральные» звуки: слова, слоги, свист. Так, различаются звуки С, Ш, Ж, 3 или даже один и тот же звук, произнесенный в разном тембре.
Лимбическая кора, входящая в эту же систему структур, является в значительной степени проекционной зоной передних ядер таламуса. Особенность состоит в том, что активность ее нейронов гасится не постепенно, а выключается сразу без привыкания, что обеспечивает еще один вид кодирования информации. Таким образом, эти структуры, как показывают проведенные исследования, работают по разному принципу и осуществляют по крайней мере три вида кодирования информации: временной, качественный и пороговый.
Оба класса структур лимбической системы функционируют в тесном взаимодействии. Если в первом фоновом цикле регистрация угасает, то второй цикл, кодирующий информацию, отключается.
Изучение лимбической системы показывает ее функциональную неоднородность, так как различные стороны процесса регистрации поступившей информации обеспечиваются различными ее образованиями.
Характерная черта дефекта памяти при поражении структур гиппокампова круга — не только распространение его на все виды материала, независимо от их модальности, но и то, что нарушение памяти касается преимущественно недавних событий, а память на отдаленные события сохраняется. Это позволяет думать, что эти образования мозга имеют значение в формировании стабильных следов памяти, так как при их повреждении не отмечается перенос следов в долговременную память. Эти данные, несомненно, свидетельствуют о том, что для образования долговременной памяти требуется какой-то дополнительный процесс, обозначаемый как процесс или фаза консолидации следа. Такие факты трудно объяснить иначе, как наличием различных механизмов для кратковременной и долговременной памяти и признать, следовательно, существование по крайней мере двух различных видов памяти.
Приведенные данные позволяют считать, что глубинные структуры мозга в области височных долей, иными словами, структуры гиппокампа, содержат механизм закрепления следов, который некоторыми авторами (в том числе отечественными неврологами JI. П. Латашом и Л. Т. Поповой) назван первичным механизмом памяти. Однако пока еще не ясно, являются ли описанные нарушения памяти первичными вследствие повреждения первичных механизмов памяти (если таковые имеются), либо вторичными вследствие повреждения структур, связанных с другими функциями мозга. На животных получены противоречивые данные.
Деление памяти на кратковременную и долговременную соответствует многим экспериментальным фактам. Так, показано, что фаза кратковременной памяти, в течение которой происходит консолидация следа памяти, легко ранимая стадия. Если в этот период мозг подвергается сильному электрическому раздражению, химическому воздействию, например, наркозу или механической травме, то консолидация нарушается и человек забывает события, непосредственно предшествовавшие этому воздействию. По мнению ряда исследователей, на все события, происходящие за 30—60 минут и более до неблагоприятного воздействия, память сохраняется, так как они прошли период консолидации. После того как завершился период консолидации, информация удерживается очень прочно. Никакие физические, химические, механические воздействия, совместимые с жизнью, уже не могут ее искоренить. Даже охлаждение организма до очень низких температур, когда исчезает совершенно электрическая активность мозга, пе приводит к забыванию событий. При восстановлении нормальной температуры сознание и память возвращаются. Это дает основание утверждать, что наряду с первой динамической фазой памяти существуют и статические ее механизмы, обеспечивающие долговременное хранение информации.
Эксперименты на животных позволили установить время, необходимое для консолидации следа памяти. С этой целью крыс обучали находить путь в лабиринте. Затем, раздражая мозг крысы электрическим током, вызывали приступ судорог. Если электрическое раздражение наносилось крысе не позже получаса после пробега по лабиринту, то память о лабиринте исчезала, наступала ретроградная амнезия и обучение надо было начинать сначала. Если электрошок проводили спустя 45—60 минут и более, то память о найденном в лабиринте пути сохранялась. Значит, воспринятая информация прошла уже период закрепления. В начале периода консолидации электросудорожный шок может искоренить информацию из мозга полностью. Но по мере удлинения времени закрепление становится все более устойчивым и уже не поддается разрушению. При этом полная амнезия, при которой больной полностью забывает все, наблюдается в пределах нескольких минут, полная неэффективность шока при интервале около одного часа, который и принимается за время консолидации следа.
После электрошока процесс консолидации восстанавливается не сразу. Поэтому исчезает память не только на события, предшествующие электрошоку, но и на события, имевшие место в течение определенного времени после электрошока, т. е. наступает период аитероградной амнезии.
Таким образом, электрошок, так же как и другие повреждающие мозг воздействия, нанесенные тотчас или вскоре после обучения новому навыку, приводит к амнезии на события, предшествующие и непосредственно следующие за электрошоком. О механизме происхождения этого амнезирующего эффекта может быть высказано предположение, что он связан с разрушением следа памяти в результате нарушения процессов его консолидации. В пользу этого предположения говорит наблюдаемая зависимость выраженности амнезирующего эффекта от периода времени между обучением и воздействием повреждающего фактора. Это основной аргумент в пользу теории консолидации.
Большинство ученых, учитывая крайнюю неустойчивость следов кратковременной памяти, полагают, что в основе их образования лежат не структурные, а нейро- динамические факторы. Скорее всего речь идет о многократной циркуляции импульсов в замкнутой системе нейронов. Мы принимаем эту гипотезу в качестве объяснения кратковременной памяти, хотя не все фактические данные с нею согласуются: результаты некоторых экспериментов показывают, что по мере приобретения навыка электрическая активность, которую можно зарегистрировать, продолжается недолго и ослабевает, однако возможно, что быстро затухающая электрическая активность вызывает ряд последовательных химических процессов, которые и поддерживают кратковременную память до тех пор, пока не возникает более стойкое запоминание. Вопрос о природе нейродинамических сдвигов, являющихся носителями следов, не может считаться в настоящее время окончательно выясненным, так как нет еще решающих экспериментальных данных.
Один из видов кратковременной памяти — оперативная память, связанная с выполнением определенной деятельности. Этот вид памяти можно проиллюстрировать множеством примеров. Так, удается удержать в памяти незнакомый номер телефона, пока мы его не набрали. Мы помним также весь перечень необходимых покупок, пока мы их не произвели, и последовательность нужных операций, пока мы не завершили действия, намеченного на данный момент. Этот вид кратковременной памяти наименее изучен. Полагают, что в основе механизмов долговременной памяти лежат какие-то структурные изменения, вероятнее всего в области межнейрональных контактов — синапсов (от греческого synapsis, что значит «застежка, связь, соприкосновение»). В настоящее время наиболее общепринято представление о наличии двух стадий фиксации в мозгу прошлого опыта: стадии нейродинамических сдвигов (кратковременная память) и стадии структурных изменений (долговременная память), являющихся следствием упомянутых нейродинамических сдвигов и представляющих собой две стадии единого процесса.
Большой интерес в этом плане представляют данные, полученные па человеке при изучении последствий сотрясения мозга. Невролог Р. Расселл из Оксфордского университета исследовал амнезии, вызываемые сотрясением мозга. Анализ большого количества случаев показал, что длительность предшествовавшего травме периода, память о котором никогда не возвращается, — постоянная ретроградная амнезия — чаще всего исчисляется всего несколькими десятками секунд. Это явление указывает, по-видимому, что след текущего события в памяти угасает очень быстро — в течение секунд, если какой-то механизм не отбирает некоторые элементы информации, чтобы зафиксировать их в кратковременной памяти. Результаты изучения ретроградной амнезии свидетельствуют о том, что для положительного действия этого механизма требуются лишь секунды. Именно перерывом процесса в результате травмы мозга и объясняется постоянная ретроградная амнезия. Согласно приведенным данным, любое впечатление внешнего мира сохраняется в мозгу несколько секунд, а затем угасает и только мозговой механизм, по-видимому, концентрирующий внимание, отбирает какую-то часть полученной информации для более длительного ее хранения. Учитывая это, можно говорить о наличии не двух, а трех стадий формирования следа памяти: стадии «мгновенной» памяти, длящейся секунды, стадии «промежуточной», или кратковременной памяти, соответствующей периоду консолидации, и долговременной памяти, длящейся, по-видимому, всю жизнь.
О чрезвычайной прочности старых следов памяти свидетельствует целый ряд фактов. Многими наблюдениями доказано, что запомнившиеся события не забываются и при определенных условиях снова могут быть извлечены из памяти. Часто мы видим во сне давно забытые лица. Оказывается, они удерживаются в мозгу и при определенном функциональном его состоянии вновь выносятся на поверхность свежими и яркими, как будто это вчерашние впечатления. Известно, что пекоторые люди в лихорадочном бреду говорят о событиях, о которых по выздоровлении не вспоминают. Удается, однако, установить, что эти высказывания в бреду являются отражением действительных событий из далекого детства или юности. В литературе описан такой случай. Английский священник посетил замок. Когда он подошел к его воротам, ему показалось, что он уже когда-то был тут и видел не только дорогу, но и ослов и людей. Через некоторое время мать сообщила, что когда ему было всего полтора года, опа в обществе друзей посетила этот замок, взяв его с собой. Некоторые из спутников закусывали на крыше над воротами, в то время как ребенок оставался внизу. И вот при втором посещении вид ворот оживил впечатления раннего детства.
Известна другая такая же история. К женщине, умиравшей в деревенском доме, привезли из Лондона совсем маленькую дочь. После короткого свидания ее снова отвезли в город. Девочка выросла. Не имея ни малейших воспоминаний о матери, уже в зрелом возрасте дочь случайно попала в комнату, где умерла ее мать, и содрогнулась. На вопрос друга она ответила, что живо припоминает, как уже однажды была в этой комнате, и помнит, что женщина, лежавшая в углу, наклонилась над ней и плакала.
Уместно вспомнить данные крупнейшего канадского нейрохирурга У. Пенфилда, о которых позднее будет сказано более подробно, о том, что при электрическом раздражении височной коры головного мозга в памяти больного могут оживать эпизоды далекого прошлого, которые воспроизводятся в таких деталях, какие кажутся просто недоступными для нормальной памяти. Эти наблюдения придают правдоподобие давно уже известным из литературы феноменальным проявлениям памяти в гипнозе.
Американский физик Д. Вулдридж рассказывает о 60-летнем каменщике, который в состояпии гипноза мог описать отдельные кирпичи в стене, выложенной им в возрасте между 20 и 30 годами. Точность описания была проверена. Кирпичи оказались там и были именно такими, как в описании.
Факты сохранения в мозгу записи прошлых событий, далеко превосходящей то, что мы можем обычно воспроизвести сознательно, могут создать впечатление, будто информация, которую мы получаем на протяжении всей нашей жизни, полностью сохраняется в памяти. Однако нет достаточных оснований считать, что сохраненная в мозгу информация приближается по своему объему к первично воспринятой. Приведенные факты свидетельствуют лишь о том, что наша память хранит больше данных по сравнению с тем, чем мы оперируем. С другой стороны, в связи с этими фактами становится очевидным, что забывание носит лишь временный характер и что наблюдаемые нередко пробелы памяти, когда не удается вспомнить нужное имя, название книги, формулу и т. д., связаны не с распадом следа памяти, а с несовершенством аппарата воспроизведения. О временном характере забывания заставляют думать и некоторые клинические факты. Так, больные с грубым нарушением памяти, будучи не в состоянии вспомнить то, что хотели запомнить, уже через 20—30 минут после ознакомления, могут сделать это спустя сутки или несколько часов. Такие факты, несомненно, свидетельствуют о наличии в мозгу долговременного следа, находящегося при обычных условиях в латентном состоянии и не поддающегося извлечению.
Наибольшим признанием в настоящее время пользуется объяснение забывания на основе феномена интерференции, сущность которого мы уже изложили в начале этой главы.
Амнезирующий эффект электросудорожного шока усматривают с позиции явлений интерференции — между запоминаемым событием и реакцией на шок. Таким же образом объясняют воздействие острой черепно-мозговой травмы — наступление ретроградной амнезии. Поскольку и при других амнестических состояниях выявлены признаки усиленной интерференции, то и суть дефекта па-’ мяти объясняют этим усилением, приводящим к ускоренному забыванию. Такой подход к сущности процесса забывания стал важным возражением против теории консолидации. Поэтому теория консолидации не пользуется общим признанием.
В настоящее время получен еще ряд экспериментальных фактов, которые полностью не укладываются в рамки теории консолидации. Было обнаружено, что амнезирующий эффект в виде ретроградной амнезии иногда наступает не сразу, а через несколько часов после травмирующего воздействия, в то время как амнезия, связанная с нарушением процесса консолидации, должна проявиться сразу же после травмы вследствие прерывания этого процесса и распада еще не успевших консолидироваться следов. Признание же фазы консолидации в процессе формирования следа памяти исключает влияние времени на возникновение амнезии.
Оказалось также, что при повторном обучении или выработке условного рефлекса, как будто бы полностью забытого вследствие воздействия электрошока, новый навык становится устойчивым к его действию, независимо от времени воздействия. Несомненно, это должно указывать на существование долговременного следа, сформировавшегося при первичном обучении, но остававшегося до повторного обучения в недоступном для произвольного воспроизведения состоянии.
Определенное значение среди описанных фактов имеет феномен «напоминания», когда действие безусловного раздражителя, например пищи, в совершенно незнакомой для животного обстановке приводит к устранению амнезии и воспроизведению навыка, заученного до электрошока. Этот факт также можно объяснить только сохранением следов ранее заученного навыка. Такого рода дан- пые получены Р. И. Кругликовым.
С позиций теории консолидации не совсем понятны и некоторые клинические факты, например, наблюдаемые при эпилептических припадках насильственные воспоминания событий, которые непосредственно предшествовали травме, обмороку и другим повреждающим мозг факторам. В связи с этим соответствующие следы не могли быть, по-видимому, закреплены и переведены в долговременную память.
Все это заставляет думать, что основу ретроградной и, может быть, антероградной амнезии составляет не разрушение следа, а какие-то другие факторы, связанные с формой хранения информации. Из этого следует, что все виды амнезии, и в первую очередь ретроградная амнезия, являются следствием нарушения процессов, ответственных за воспроизведение образовавшегося следа. Это должно означать, что ретроградная амнезия после амнезиру- ющих воздействий обусловлена не разрушением формирующегося следа, а повреждением аппаратов его воспроизведения, либо переводом следа в подпороговое, недоступное для сознательного воспроизведения, состояние.
Приведенные факты дают основание считать, что формирование следа памяти является почти мгновенным процессом, и, следовательно, понятие кратковременная память, отражающее согласно теории консолидации период закрепления следа, должно получить другую интерпретацию. Имеющиеся наблюдения заставляют считать недоказанным наличие фазы консолидации следа, и в таком случае к кратковременной памяти может быть отнесена только «мгновенная» память, существующая в течение секунд и не имеющая отношения к закреплению следа в долговременной памяти. Однако, несмотря на приведенные возражения, гипотеза консолидации следа пока лучше других согласуется с основными экспериментальными и клиническими фактами.
Где же хранятся следы памяти?
Первые попытки выяснить это были направлены на изучение эффектов, возникающих при удалении различных областей коры головного мозга, поскольку функциональное значение корковых структур связывалось со сложными психическими функциями, прежде всего с мыслительными процессами и анализом поступающей информации. Но ни очаговые поражения корковых структур, достаточно часто наблюдаемые в клинической практике, ни многочисленные перерезки и удаления различных корковых зон, проведенные американским ученым Лешли «в погоне за энграмой» (в поисках следа памяти), не приводили к выраженному дефекту памяти. В связи с отрицательными результатами многолетних опытов Лешли пришел к выводу о невозможности точной локализации следов памяти в каких-нибудь изолированных участках коры и о равнозначности в этом отношении всех ее областей.
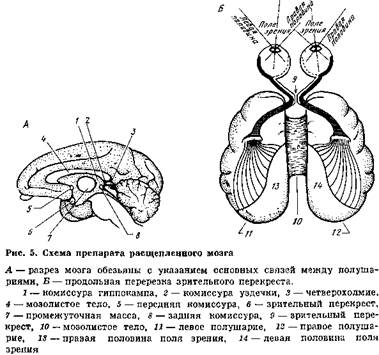
Опыты зарубежных ученых Р. Сперри и Я. Буреша, проведенные на животных с так называемым «расщепленным мозгом», т. е. в условиях полного хирургического разделения полушарий путем срединной перерезки, в некоторой степени приблизили нас к решению вопроса о месте хранения следов, связанных с навыками различения. Экспериментаторы пришли к выводу, что такого рода следы памяти прежде всего находятся в коре больших полушарий. Опыты проводились на кошках, которых предварительно обучали некоторым видам зрительного различения. В камере с двумя дверцами на одной вывешивались круг, а на другой — квадрат. Кошку обучали открывать дверцу только с кругом. Существенную роль в опыте играла повязка, позволявшая подопытному животному видеть только одним глазом. Закрыв кошке правый глаз, ее обучали различать эти две фигуры. Как только животное обучалось делать правильный выбор, повязку перемещали на другой глаз. В опытах с нормальной кошкой не было различия в результатах, хотя при обучении использовался только один глаз. Затем Сперри отделял у кошек одно полушарие от другого путем перерезки связей, проходящих через мозолистое тело и некоторые другие группы волокон. Одновременно он перерезал зрительный перекрест, чтобы обеспечить поступление информации из правого глаза только в правое полушарие, а из левого глаза — в левое.
На этот раз результаты поразительно отличались от прежних. Кошка с расщепленным, или раздвоенным мозгом, обученная почти безошибочно различать фигуры с помощью левого глаза, после перемещепия повязки вела себя так, как будто попала в эту ситуацию впервые. Следовательно, никакого переноса навыка из одного полушария в другое не происходило. Животных с раздвоенным мозгом удавалось обучить даже одновременно двум противоположным навыкам, предъявляя разные фигуры правому и левому глазу и, следовательно, адресуя их в разные полушария мозга. Аналогичные исследования проводились с введением информации не только через зрительные пути, но и через слуховые, обонятельные и другие анализаторы. Опыты ставили и на других животных, в том числе па обезьянах.
Во всех опытах на животных с расщепленным мозгом выявлялась независимая работа каждого полушария. Оказалось, что опыт, приобретенный до операции, проявился в деятельности обоих полушарий, тогда как опыт, полученный после перерезки в условиях изолированного обучения одного полушария, выявился только соответствующим полушарием (рис. 5). Полученные результаты могут быть поняты только в том случае, если мы предположим, что благодаря связи между полушариями через мозолистое тело происходит либо дублирование, либо просто сравнение следов памяти, поступающих в кору одного полушария, со следами, хранящимися в коре другого. Опыты Сперри и его коллег убедительно показали, что благодаря связи между полушариями через мозолистое тело действительно происходит дублирование следов в обоих полушариях, так как в условиях «расщепленного мозга» выработанные ранее следы были представлены в каждом полушарии.
Последующие опыты этой школы исследователей с удалением значительных областей мозговой коры после обу-
|
|
чения показали, в какой именно части ее хранятся эти следы. Так, кошек с «расщепленным мозгом» обучали навыку различения, заключавшемуся в том, что они должны были нажимать левой передней лапой на один из двух рычагов — с гладкой и шероховатой поверхностью. После этого у них удаляли всю кору правого полушария, связанного с левой половиной тела, кроме чувствительной и двигательных ее зон, необходимых для осуществления этого навыка. Оказалось, что оставшаяся кора способна обеспечить отличное выполнение усвоенного акта; животные могли также обучаться новым аналогичным навыкам с помощью той же левой лапы. Таким путем Сперри удалось ограничить возможную локализацию следов памяти для этих навыков определенной зоной — в данном случае сенсо-моторной, чувствительно-двигательной корой
соответствующего полушария, связанной как с восприятием чувствительных импульсов от левых конечностей, так и осуществлением двигательной программы выработанного навыка.
К таким же выводам пришел чешский нейрофизиолог Я. Буреш, пользуясь совершенно другим методом исследования. Буреш назвал свой метод обратимой декортикацией. Смачивая обнаженный мозг раствором хлористого калия, он вызывал явление, называемое распространяющейся депрессией, которое приводило к обратимой инактивации, т. е. выключению обработанного таким путем участка коры. С помощью этого метода Бурешу удавалось на протяжении нескольких часов выключать целое полушарие мозга. Так, при воздействии хлористым калием на кору обоих полушарий животное теряло способность решать даже простые задачи. Но при выключении коры лишь одного полушария оно сохраняло нормальные способности. У крыс вырабатывались простые навыки в период, когда одно полушарие было отключено. На следующий день инактивировали другое полушарие и пытались заставить животное выполнять усвоенную задачу. Оказалось, что никакого переноса следов памяти не происходило, и обучение надо было начинать сызнова.
Во время обучения перенос следов происходит лишь тогда, когда животное несколько раз повторяет усвоенную задачу. Только в этом случае при последующем выключении «обученного» полушария второе полушарие тоже оказывается «обученным».
Из опытов Буреша был сделан вывод, что перенос навыков в нормально функционирующем мозгу не может совершаться вне обучения, а осуществляется только благодаря переносу навыков во время «повторений».
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|