- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Мембраналық моделдер
Мембраналық моделдер
Биологиялық мембрананың құрылымының алғашқы моделі 1902ж. ұсынылған. Мембрана арқылы липидтерде жақсы еритін заттар тез өтетінін байқап, биологиялық мембрана фосфолипидтің жұқа қабатынан тұратындығы тұжырымдалған. Фосфолипид молекуласы су-ауа (су-май) бөлігінің шекарасында бір қабат құрып, яғни гидрофильді (полярлы) бастары сумен, ал гидрофобты «құйрықтары» бастары ауада (майда) орналасқан. Сондықтан, биологиялық мембрананың бірінші моделі -липидтердің моноқабатына (4, А сурет) негізделген. 1925 ж. Гортер және Грендель гемолизденген эритроциттерден липидтерді ацетон арқылы бөліп алып, оларды су бетіне құйып, пайда болған бір қабаттан тұратын мономолекулалар пленкасының ауданын анықтаған. Эритроциттерден бөлініп алынған бір қабат болып орналасқан липидтер бетінің ауданы сол эритроциттердің ауданынан екі есе көп болғандығы анықталған. Осы зерттеу нәтижесінде мембранадағы липидтер бимолекулалық екі қабат түрінде орналасқан деген тұжырым жасалынды (4, Б сурет).
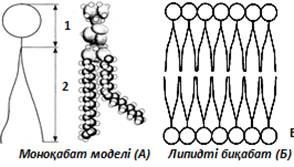
4 сурет. Фосфолипид схемасы:
А)1 - полярлы (гидрофильді) бөлігі; 2 - полярсыз (гидрофобты) бөлігі
Б) Бимолекулалы фосфолипидті қабат
Бұл тұжырымның дұрыстығы биологиялық мембрананың электрлік параметрлерін өлшеу барысында дәлелденді (Коул және Кертис, 1935 ж.): биомембрананың электрлік кедергісі ≈107Ом×м2 мен электрлік сиымдылығының ~0,5×10-2Ф/м2 үлкен болатындығын анықтады.
Жоғарыда мембрананың құрыламында айтылғандай, биологиялық мембрана ақуыз молекуласынан тұрады. Мысалы, тәжірибелер нәтижесінде жасушалық мембрананың беттік керілуін өлшеу кезінде өлшенген беттік керілу параметрлері липид-су (10-2Н/м жуық) бөлігінің шекарасына қарағанда ақуыз-су бөлігінің шекарасының беттік керілу коэффициентіне (10-4Н/м жуық) жақын. Осы қарама-қайшылықты 1935ж. Даниэль мен Давсонның тәжірибелік нәтижесі жойып, биологиялық мембрана құрылымының мембранологияда 40жыл үстем еткен «бутербродтық» моделін ұсынды. Бұл моделде мембрана – үшқабаттан тұрады. Модель бойынша фосфолипидтер молекулалары мембрана бетінде перпендикуляр түрде екі қатар болып орналасқан. Липид молекулалары гидрофильді жағымен мембрананың сыртында, гидрофобты жағымен оның ішіне қарай бағытталып орналасады. Мембрананың екі қатар орналасқан фосфолипид молекулаларының полярлы топтарына ақуыз молекулалары жабысып орналасады, соның нәтижесінде мембранада иілгіштік, механикалық әсерлерге беріктілік, беттік керілу төмен қасиет пайда болады.
Жалпақ биқабатты липидті мембраналар (БЛМ). Мембрананың екінші моделі -жалпақ биқабат.Бұндай мембраналарды су құйылған пластикалық (мысалы, фторопласт) материалдардың диаметрі 1мм жуық кішкене тесіктерінде алынады, яғни липидтер биқабатты құрылымдарды (қара пленкаларды) құрайды. Бұл құбылыс алғашқы рет О. Мюллермен зерттелген. Ол екі су фазаны шектейтін, ауданы 0,5-5,0 мм2 тефлондық қалқаның кішкене тесіктерінде мидың фосфолипидтерінен биқабатты липидті мембраналарды (БЛМ) алған. БЛМ қалыптасу процесі сұйық көмірсуларда ерітілген липидті тефлондық стақанға жағудан басталады (5 сурет).
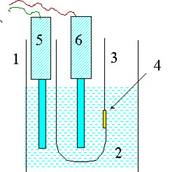
5 сурет. Бимолекулалық липидті мембраналарды дайындау. (1) шыны стақанға (2) электролит ерітіндіні орнатады және (4) тесігі бар тефлонды ыдысты ішіне малтырады.
Тесікте БЛМ қалыптасады (6сурет). Капиллярмен тесікке фосфолипидтің кішкене тамшысын орнатады. Фосфолипидтің молекулалары келесі түрде орналасады. Полярлық бастары су ортасына қарайды, ал гидрофобты құйрықтары тамшының ішіне тығылады. Біртіндеп тамшыдан ерітуші кетеді да тамшы липидті пленкаға айналады.
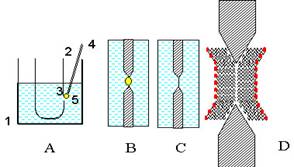
6 сурет. БЛМ тефлон ыдыстың қабырғасында пайда болуы. А - (4) капиллярдың көмегімен (3) ыдыстың қабырғасындағы тесікке (5) гептандағы фосфолипид ерітіндісінің тамшысын енгізу.
Пленканың мінезін анықтайтын басты күштері - (σ) фазааралық (үстінгі) тұтқырлық және ван-дер ваальс күштері.
Алғашқы уақытта липидті пленканың қалындығы 100 нм-ден жоғары болады (7 сурет).
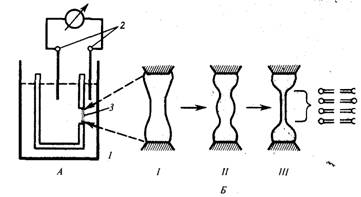
7 сурет. БЛП қалыптасу кезеңдері (I-қалың мембрана, II - дөңес линзатәрізді мембрана,III - БЛМ).
Пленканың жіңішкеруін және БЛМ құрылуын шағылысу сәулесі арқылы байқауға болады. Алғашқы кезенде, пленка қалың болғанда, ол кәдімгі макродене сияқты көрінеді. Пленканың қалыңдығы түскен жарықтың толқын ұзындығына жақындағанда, сәулелердің интерференциясы (Ньютон сақиналары) пайда болады. Мембрананың үстінде түсті оюлар пайда болады. Биқабатты липидті құрылымдар боялған түсте қара болып көрінеді, сондықтан, олар қара пленка деп аталып кетті. Ондай пленкалардың төмен шағылу қабілеті келесі фактпен байланысты. Пленканың алдынғы және артқы жазықтықтарынан шағылған сәулелер қарсы фазада тұрады да, бір-бірін сөндіреді.Қарайғаннан кейін, 15-20 минут арасында үстінгі тұтқырлық төмендейді де, БЛМ-ның электрік сыйымдылығы кейбір стационарлы мәндерге жеткенше өседі (8 сурет).
8 сурет. Жалпақ биқабатты липидті мембрананың құрылуы
а) қалыптасу процесі, б) қалыптасқан
БЛМ-дың электрлік сипаттамалары және басқа физикалық-химиялық қасиеттері биологиялық мембраналардың қасиеттеріне ұқсас. Бірақ олардың метаболизмдік белсенділігі өте төмен.Жалпақ липидті мембраналар, липосомалармен қатар мембраналардың электрлік қасиеттерін, өткізгіштігін және басқа қасиеттерін зерттеу үшін қолданады. Моделді мембраналардың көмегімен баръерлік, селекивтік (су үшін жоғары, ал иондар үшін төмен деңгейдегі өткізгіштік) қасиеттерін зерттеуге болады. Моделдік мембранаға тасымалдаушы-молекулаларды енгізіп, биологиялық тасымалдауды моделдеуге болады.Мембрананың құрылымының және функцияларының зерттелуі, табыстар мен жетістіктер ең алдымен моделді тәжірибелерден алынған мәліметтерде негізделеді.
Липосомалар.Липосомаларды немесе фосфолипидті везикулаларды липидтерді суға шашырату арқылы алады. Мембрананың үшінші моделі – сфералық бірқабатты (бірламеллярлы) липосома. Олардың полярлық топтары суда, ал гидрофобты топтары ауада орналасады, мономолекулярлы қабатты құрып, жайылады (9 сурет).Олар биқабатты мембранадан тұратын, ұсақ көпіршіктер (везикулалар) түрінде болады. Липосомалар толық ақуыз молекуласынан айырылған, биологиялық мембрана болып табылады. Жоғарыда айтылғандай, липосомалар алғашфосфолипидтен алынған. Жасушадағы фосфолипидтің қызметі – мембрананың компоненттерін құру. Олар өздерінің химиялық құрамы жағынан амфифилді қосылыстар деп аталатын топқа жатады. "Липосома" және "липидті везикулалар" сөзі синоним ретінде жиі қолданылады. Бірақ тарихи тұрғыда липосомалар алғаш суда ісінген фосфолипидтерді механикалық шашыратуда қалыптасқан бөлшектер деп қарастырады. Бұл бөлшектер, бірнеше ондаған, тіпті жүздеген сулы аралықпен бөлінеген липидті биқабаттан тұратын көп қабатты мультиламеллярлы везикулалар (МЛВ) және өте кішкентай бір липидті биқабаттан тұратын моноламеллярлы везикулалар (ММВ) деп аталады. Липосомалардың сыртқы пішіні ылғи да глобулярлы бөлшектер тәрізді емес. Кейде олар дисктәрізді (дискома деп аталып) формалы немесе өте ұзын жіңішке түтікше тәрізді формаға ие болып, тубулярлық липосомалар деп аталады.

|
| 9 сурет. Липосомалар: а - бірқабатты; б – көпқабатты. |
Бірқабатты липосоманы әртүрлі әдіспен алуға болады, мысалы, ультрадыбыстың әсерінен көпқабатты липосома суспензиясынан. Осы әдіспен алынған бірқабатты липосоманың диаметрі 25-30 нм құрайды. Моноламеллярлық липосомаларды көптеген мидико-биологиялық зерттеулерде қолданады. Бірақ, олардың кішкентай көлемі мен осмостық пассивтілігімен байланысты зерттеулерге кедергі жасайды. Қазіргі заманда диаметрі 400 нм. астам үлкен моноламелярлы липосомаларды жасау әдістемелері дамыған.Әрекеттесулердің мінезіне сәйкес не мульти, не моноламелярлы липосомалар пайда болады. Қарапайым механикалық әрекеттесулердің нәтижесінде көпқабатты бөліктер пайда болады (диаметрі бірнеше микрометр). Көпқабатты липосоманың жеке бимолекулалық қабаттары су ортасымен ерекшеленеді. Липидті қабаттардың қалындығы 6,5 - 7,5 нм, ал ара қашықтық - 1,5 - 2 нм. Көпқабатты липосоманың диаметрі 60 нм мен 400 нм арасында болады.Бұл бөліктерде липидті биқабаттар ішкі фазаны сыртқы ерітіндіден бөледі. Бұнымен байланысты мультиламелярлы липосомаларды липидті биқабаттың барьерлік қызметін зерттеу үшін қолданады.Мультиламелярлы липосомалар осмостық белсенді, сыртқы ортаның осмостық қасиеттері өзгергенде олардың көлемі өзгереді.Липосома көмегімен мембрана арқылы зат тасымалдануының негізгі заңдылықтары анықталған, мембрананың қызмет атқаруындағы фазалық ауысулардың маңыздылығы көрсетіліп, липидті биқабаттың молекулярлық параметрлері анықталды және оның динамикалық сипаты, жекелей мембрананың ақуыздары мен толық ақуыздар ансамблдері процестері зерттелген.Соңғы кезде липосомалар мен везикулаларға көптеген фундаменталды ғылымдардың өкілдері қызығушылық танытуда. Математиктерді липидті биқабаттың серпімді құрылымына байланысты үш өлшемді кеңістіктегі екі өлшемді үстілік топологиясы жағдайлары қызықтырады. Физиктерді липосомалар фракталды жүйе ретінде қызықтыруда, яғни агрегация кезіндегі оның ерекшелігі және морфологиялық ауысулардың сипаты. Химиктер үшін биқабатты везикулалар белгілі бір ортада химиялық реакциялардың жүруіне мүмкіндік беретін микрореакторлар ретінде қызықтыруда.
Липосомаларды медицинада кеңінен қолданады. Мысалы, липосомаларды дәрілерді ағзалар мен тіндерге жеткізу үшін фосфолипидті микрокапсула ретінде қолданады. Липосомалар табиғи липидтерден алынғандықтан олар токсинді емес (липидті дұрыс таңдау барысында), кері иммундық реакциялар туғызбайды, организмде толық сіңеді, кейбір биологиялық тосқауылдарды еңсеруге қабілетті. Мысалы, липосомадағы инсулин асқорыту ферментінің әсерінен сақталған. Қазіргі кезде ісіктер, атеросклероз, фермент жеткіліксіздігі липосомальді терапия әдістері дамуда. Бірақ липосомды терапевтикалық қолдану жағдайы ойлағандай оңай емес. Қанда липосома айтарлықтай тұрақсыз және бауырдағы, көкбауырдағы, жұлындағы магрофагтармен бірге қан ағысымен тез шығып кететін болып шықты. 1987 жылы екі атақты косметикалық компаниялардың лабораториялық зерттеулері жемісінің жаңа өнімін ойлап тапты. Бұл "Кристиан Диор"фирмасының "Каптюр"липосомалық гелі және "Л'Ореаль"фирмасының "Ниосома" теріге арналған кремі.
Липосомалардағы дәрілік препараттарды ауырған органға немесе ауырған бөлікке (жекелей, жүректің зақымдалған бөлігіне) тура бағытты жеткізу мүмкіншілігі зерттелуде. Бұл үшін липосомаға ақуыздық молекула – антидене қосылады.
Протеолипосомалар.Көптеген мембраналық ақуыздарды жасанды везикулалық мембраналардың құрамына енгізуге болады. Ондай комбинативті жүйелер протеолипосомалар деп аталады. Ақуыздарды енгізу тиімділігі мембраналардың липидті құрамынан, рН, тұздық құрамынан, температурадан және т.б. тәуелді. Егер де детергенттер қосылса, ақуыз молекулалардың ену тиімділігі өседі. Протеолипосомаларды медицинада қолданады. Протеолипосомаларды адам организміне енгізуде ол жасушалық мембранамен сіңіріледі. Биомембранамен байланысып, липосома олардың құрамын өзгертеді. Плазмалеманың бір жасушасына 300 млн. липид молекуласын енгізіп, жасушалық мембрананың негізгі құрылымын (тұтқырлық, өткізгіштік, ферметтік және иммундық қабілеттілігі) өзгертуге болады. Мұның маңызды жетістігі сол, протеолипосомалар организмнің барлық жасушаларында емес, сол емге зәру болып тұрған жерге таралады. Жасушалық деңгейдедегі тура бағытталған терапия әртүрлі әдістермен, жекелей, липосомаға ақуыздық компоненттерді енгізумен іске асырылады. Протеолипосомалар дәрілік құрал ретінде токсинді емес, организмде толық сіңеді, кейбір биологиялық тосқауылдарды еңсеруге қабілетті.
МЕМБРАНАНЫҢ ҚҰРЫЛЫМЫ ТУРАЛЫ ҚАЗІРГІ УАҚЫТТА ҚОЛДАНЫЛАТЫН ҰҒЫМДАР. СИНГЕР МЕН НИКОЛЬСОННЫҢ СҰЙЫҚ КРИСТАЛЛ МОДЕЛІ.
Алынған физикалық және химиялық зерттеулер нәтижесінің жиынтығы жаңа сұйық мозайкалық моделді ұсынуға мүмкіндік берді (Сингер және Никольсон, 1972 ж.). Биологиялық мембрананың негізін ақуыздармен оюланған, фосфолипидтердің екі қабаты құрайды. Биологиялық мембранада фосфолипид пен ақуыздан басқа да химиялық қосылыстар бар. Жануарлар жасушасының мембранасында холестерин көп. Мембранада гликолипидтер, гликопротеидтер бар.
Моделде биологиялық мембрана липидтері сұйық күйде болады, липидте ақуыздары жүзіп жүреді, бірақ ақуыздар липидте үнемі бос жүзбейді, ішкі (цитоплазмалық) жасушалық құрылымға зәкірленеді. Бұндай құрылымға микрофиламенттер және микротүтікшелер жатады (13 сурет).
Микротүтікшелер – жасуша қызметінде маңызды роль атқаратын диаметрі 300 нм болатын, іші қуыс цилиндрлі ерекше ақуыздан (тубулин) тұрады.
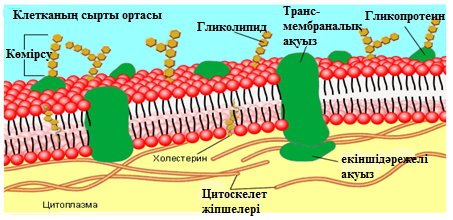
10 сурет. Плазмалық мембрананың сұйық-мозаикалық үлгісі.
Сонымен қатар, мембранадағы липидтердің барлығы биқабат принципі бойынша орналаспаған. Физикалық әдістер негізінде, мембрананың орналасатын бөліктері бар екендігін көрсетті. Мембрананың құрылысының, мембрана ақуыздарының және басқа да заттардың күрделі химиялық құрамын зерттеумен биохимия айналысады. Биофизика мембрананың негізгі құрылымын, нақтырақ фосфолипидті молекуланың биқабатын зерттейді.
Лецитин фосфолипидінің молекуласы полярлы бастардан (фосфор қышқылының) және ұзын полярсыз құйрықтарынан (май қышқылдарының қалдықтары) тұрады. Лецитиннің фосфолипидті молекуласының бастары бір-бірінен кішкене қашықта орналасқан екі зарядталған топтан тұрады. Абсолютті шама бойынша тең әртүрлі екі заряд электрлік диполь қалыптастырады.
Мембранада фосфолипидтер әртүрлі болады. Мысалы, эритроцит мембранасында олардың түрі 20 жуық.
МЕМБРАНАНЫҢ ФИЗИКАЛЫҚ ҚҰРЫЛЫМЫ
Мембрананың қызмет атқаруы липидті биқабаттың микротұтқырлығына және мембранадағы фосфолипид қозғалғыштығына, мембрана липидтерінің фазалық жағдайына байланысты. Жасуша физиологиясында биомембраналардың фазалық ауысулары маңызды роль атқарады. Биологиялық мембрананың липидті фазасы физиологиялық жағдайдағы (температурада, қысымда, қоршаған ортаның химиялық құрамында) агрегат күйі сұйық. Бұл флюоресцентті анализ, электронды парамагнитті резонанс (ЭПР), ядролық магнитті резонанс (ЯМР) әдістері арқылы дәлелденген.
Заттар әртүрлі температураға, қысымға, химиялық компонентер концентрациясына байланысты түрлі физикалық күйде болуы мүмкін, мысалы, газ, сұйық, қатты, плазмалық күйінде болады. Мембрананың липидты биқабаты физиологиялық жағдайда – сұйық, сонымен қатар, мембрана молекулалары ретсіз емес, олардың орналасуында реттілік байқалады. Фосфолипидті молекула екі қабат құрып, ал олардың гидрофобты құйрықтары шамамен бір-біріне қарама-қарсы орналасады. Полярлы гидрофильді бастарының бағытында да белгілі рет бар. Бұндай молекулалардың орналасуы мен бағытының өзара қашықтық реттілігі бар, бірақ агрегат жағдайы сұйық физикалық күйін сұйықкристалды деп атайды. Сұйықкристалды күйдің құрылымы әртүрлі болуы мүмкін (14, б, в, г сурет): нематикалық (жіңішке жіп тәрізді), ұзын молекулалары бір-біріне қарама-қарсы бағытталған; смектикалық (сабынтәрізді) – молекулалар бір-біріне қарама-қарсы, қабат болып орналасқан; холестикалық – бір жазықта молекулалар бір-біріне қарама-қарсы орналасқан, бірақ әртүрлі жазықта молекулалар бағыты әртүрлі. Биологиялық мембрананың липидті биқабаты смектикалық сұйықкристалды күйге сәйкес келеді.

11 сурет. Молекуланың аморфты (а) және сұйықкристалдыорналасуы (б-нематикалық фаза, в-смектикалық фаза, г-холестикалық фаза)
Сұйықкристалды құрылым температураның, қысымның, химиялық құрылымның, электрлік өрістің өзгерісіне өте сезімтал. Бұл мембрананың липидті биқабатының динамикалығын анықтайды, яғни олардың құрылымының түрлі өзгерістерге, тіпті ішкі химиялық құрамның немесе жағдайдың кішкене өзгерісіне де басқа фазалық күйге (мысалы, газ күйінен сұйық күйге, сұйық күйден қаттыға, бір кристалдық модификациясынан басқасына) ауысуы.
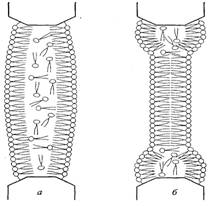
Биологиялық мембрананың липидті бөлігі белгілі бір температурада фазалық көшулерге бейім. Рентгенқұрылымдық анализ, радиоспектроскопиялық, флюоресценттік анализ, инфрақызыл спекроскопиялық, және басқа да физикалық зерттелулер нәтижесі бойынша фосфолипидті мембранада температура төмендеген кезде сұйықкристалды күйден гелдік күйге көшеді, бұны кейде қаттыкристалды деп те атайды (15 сурет).
12 сурет. Температура өзгеруіне байланысты мембрана құрылымының өзгеруі
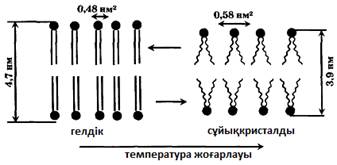
Гелдік күйде моекулалар сұйықкристалдыға қарағанда неғұрлым ретті орналасқан. Барлық фосфолипидті молекулалардың гидрофобты құйрықтары гелдік – фазада қатаң түрде бір-біріне қарама-қарсы толық созыла орналасады (толық транс-конформацияға ие). Сұйық кристалда жылу қозғалысының арқасында транс-гош-көшулері, молекула құйрықтары иіледі, сөйтіп, жекелеген жерлерде олардың бір-біріне қарама-қарсылығы бұзылады, әсіресе мембрана орталығында. Сондықтан гель-фазада мембрананың қалыңдығы сұйық кристалдыққа қарағанда көбірек (15 сурет). Бірақ, қатты күйден сұйық күйге көшу кезінде біршама көлемі ұлғаяды, себебі мембрананың ауданы бір молекулаға келетіні 0,48 нм2-ден 0,58 нм2-ге дейін ұлғаяды. Сонымен, қаттыкристалды күйге неғұрлым реттілік тән, сұйық кристалдыққа қарағанда, ол аз энтропиямен сипатталады. Мембрана қалыпты функционалдық жағдайда сұйықкристалды күйде болуы тиіс. Қоршаған ортаның температура төмендігінің ұзақтығы кезінде тірі жүйеде фазалық көшуде температураның төмендеуін қамтамасыз ететін, мембрананың химиялық құрамының адаптациялық өзгеруі байқалады. Фазалық көшуде температураның төмендеуі май-қышқылдарының құйрықтарында қанықпаған байланыстар санының ұлғаюына байланысты жүреді.
Мембрана липидтерінің химиялық құрамына байланысты фазалық көшу температурасы гель-сұйықкристал -20°С –тан (мембрананың қанықпаған липидтері үшін) +60°С –қа дейін (қаныққан липидтер үшін) өзгереді. Мембраналық фазалар арсындағы молекулалық негіздің әртүрлілігін май-қышқылы тізбектерінің конформациясы құрайды.
Сұйықкристалды күйде жеке май-қышқылды тізбек С-С байланысы айналасындағы түрленулерден әртүрлі конфигурацияларға айналуы мүмкін. Липидті биқабатта молекулалар тығыздығына байланысты қалыпты жағдайда көмірсу тізбектерінің екі жалпақ конформациясы – транс – және – цис түзіледі (16,17 сурет), бірақ аралық та бар (16, в сурет).
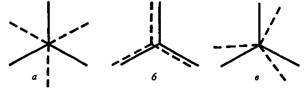
13 сурет. Этанда СН-байланыстардың транс- (а), цис- (б) және аралық (в) конформациялары
Қатты күйде фосфолипидтің барлық молекулаларының майқышқылдарының көмірсу тізбектері транс – конформацияға ие, бұл олардың билипидті мембранада қозғалғыштығын шектейді. Билипидті мембрана сұйық күйде май-қышқылдары тізбектерінің жылу қозғалысы транс-гош көшулермен жүреді. Биқабатта гош-конформация жанында қуыстар қалыптасып, биқабат қуысында - «кинктер» (ағыл. kink — ілмек) құрылуы мүмкін (18 сурет).
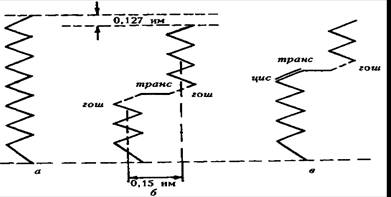
14 сурет. Транс-конфигурациядағы (а), гош-транс-гош-конфигурациядағы (б), цис-транс-гош-конфигурациядағы (в) көмірсу тізбектері
Ілмек гипотезасы (кинк) сұйыққкристалды тізбектердің фазалық көшулер кезінде еруі айналу изомеризациясына негізделген деп тұжырымдайды. Транс – неғұрлым төмен энергияға, ал сұйыққкристалды цис-конфигурация – жоғарғыға ие болып келеді. Кинктің қалыптасуы тізбек ұзындығының 0,13нм қысқаруымен сипатталады. Қазіргі кездегі тұжырым бойынша кинктер құрылымы диффузияға қабілетті (18 сурет).
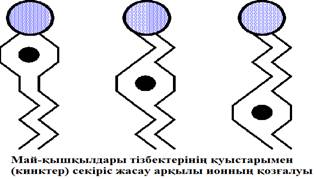
15 сурет. Мембрананың липидті биқабатында иондардың кинктер арқылы қозғалысы.
Мембранада молекула аралық байланыстар липид-липидті, липид-ақуызды және ақуыз-ақуыздық өзара әрекеттесу арқылы жүзеге асады.
Липид-липидті әрекеттесулер мыналарға байланысты:
1) электростатикалық күш энергиясына;
2) мембранада фосфолипидте «бастары» және «құйрықтарында» факторлар локализациясы;
3) сутектік байланыстардың қалыптасуы, энергиялық гидратациясы;
4) фосфолипидті доменнің қалыптасуы;
Липид-ақуыздық әрекеттесулер анықталады:
1) моноқабат үстіндегі ақуыз бен липидтің электростатикалық және сорбциялық әрекеттесуімен;
2) Билипидті мембранада ішкі мембраналық орналасумен және ақуыз бен липидтің (аннулярлы қабат) әрекеттесуімен.
3) Ақуыз-ақуыздық әрекеттесулер ақуыз молекулаларының (кластерлер қалыптасуы) мембрана ішілік және үстілік бағытына негізделген.
Биологиялық мембрананың негізгі қызметінің біреуі – бұл тасымалдау болып табылады. Мембрана арқылы тасымалданудың 4 түрі белгілі, олардың екеуі белсенді және екеуі белсенсіз. Белсенді тасымалдану, энергияны қажет етеді, ал белсенсіз тасымалдану, энергияны қажет етпейді. Плазмалық мембрана арқылы тасымалданудың қарапайым түрі: осмос және су диффузиясы.
Белсенсіз тасымалданудың басқа түрі – градиентті концентрациялы диффузия болып саналады, яғни жоғары концентрация аймағынан төменгі концентрация аймағына. Осы тасымалдану түрімен клеткаға кіші молекулалар (салмағы 150 Да дан көп емес) енеді, және де оттегі, көмірқышқыл газ және азот енеді. Бұл заттар туннелді мембраналық белоктар арқылы өтеді. Мұндай молекулаларда арнайы типті заттарды өткізетін канал болады. Белсенсіз диффузия зарядсыз иондар немесе молекулаларға тән қасиет. Белсенсіз диффузияның тағы ьір түрі липидті биқабатта заттардың ыдырауы болып саналады. Клеткаға осылай спирттер глицерол енеді.
Жеңілденген диффузия – бұл кезде клеткаға мембраналық барьер арқылы заттардың өтуіне, арнайы тасымалдаушы белоктар немесе өткізгіш белоктар көмектеседі. Бұл процесс электрохимиялық градиентпен жүреді. Өткізгіш белок, мембрананың бір жағында заттармен бірігеді де, екінші жағында оны береді. Мысалы: эритроциттерге глюкозаны тасымалдау.Тасымалдаушы белоктардың жұмыс істеу принциптері. Белок молекуласында конформациялық қозғаушыға байланысты канал түзіледі немесе «понг – пинг» механизмі деп аталады. Понг кезінде тасымалдаушы белок қосылатын бөлік сыртқа ашылады, ал ПИНГ кезінде белоктың конформациялық өзгеруіне байланысты бөлік жабылады және клетканың ішіне ашылады.
Белсенді тасымалдану кезінде энергия жұмсалады, заттарды электрохимиялық градиентке қарсы тасымалдайды. Мысалы, натрий, калий және хлор иондары. Қан плазмасында бұлардың құрамы эритроциттерге қарағанда біраз айырмашылығы бар. Соңғы зерттеулердің көрсетуі бойынша, клеткада белсенді натрий насосы жүреді, клеткадан натрий ионын шығарады. Көбінесе ол калий насосымен байланысты, клетка ішіне калий ионын енгізеді. Бұндай біріккен насосты калий, натрий насосы деп деп атаймыз. Насос – бұл мембрананы тесіп өтетін белок. Мембрананың ішкі жағынан бұған натрий ионы және АТФ жақындаса, ал сыртқы жағынан калий ионы жақындайды. Иондардың алмасуы белоктың конформациялық өзгеруіне нәтижесінде іске асады.
Тасымалдаушы белоктардың жұмыс істеу әдістері:
Тасымалдаушы белоктардың жұсым істеу әдістері 3 топқа бөлінеді: унипорт, симпорт және антипорт. Унипорт кезінде белок заттарды мембрана арқылы тасымалдайды, симпорт кезінде заттарды немесе ионды тасымалдау, басқада иондармен тығыз байланысты. Симпортқа мысал ретінде клеткаға глюкозаны өткізу, натрийдің өтуімен тығыз байланысты. Антипорт кезінде біруақытта, бір өнім клеткаға еніп жатса, екіншісі клеткадан шығып жатады. Мысалы натрий, калий насосы. Тасымалдаушы белоктардың жұмыс істеу принципі понг – пинг.Плазмалық мембрана арқылы макромолекулалар да тасымалданады. Клетканың үлкен молекуланы қоршап алуы, эндоцитоз, ал осы молекуланы клеткадан шығару экзоцитоз деп аталады. Осы тасымалдау түрлеріне ортақ нәрсе, ол тасымалдайтын заттарын плазмалық мембранамен қоршайды және көпіршік немесе везикула түрде болады. Везикуланың пайда болу механизмі және оның келешектегі тағдыры эндоцитоз типіне байланысты.
Эндоцитоз - екі топқа бөлінеді: фагоцитоз және пиноцитоз. Пиноцитоз құбылысы барлық клеткаға тән қасиет болып саналады. Бұл кезде клеткаға сұйық және кішігірім түйіршіктер енеді.
Фагоцитоз кезінде клеткаға үлкен бөлшектер енеді (жұтылады): вирустар, бактериялар, клеткалар немесе олардың сынықтары. Фагоцитоз арнаулы макрофагтар және гранулоциттар арқылы іске асады.
Эндоцитоз механизмі. Плазмалық мембранада инвагинация немесе төмпешік пайда болады. Ол қолбаға ұқсас, эноцитозды везикула деп аталады. Везикула мойыны қосылып мембранадан үзіледі. Везикула келешекте Гольджи комплексіне немесе лизосомаларға қарай тасымалдануы мүмкін, олармен қосылып, екінші лизосомалар немесе фаголизосомалар пайда болады.
Эндоцитоз - екі топқа бөлінеді: фагоцитоз және пиноцитоз. Пиноцитоз құбылысы барлық клеткаға тән қасиет болып саналады. Бұл кезде клеткаға сұйық және кішігірім түйіршіктер енеді.
Фагоцитоз кезінде клеткаға үлкен бөлшектер енеді (жұтылады): вирустар, бактериялар, клеткалар немесе олардың сынықтары. Фагоцитоз арнаулы макрофагтар және гранулоциттар арқылы іске асады.
Эндоцитоз механизмі. Плазмалық мембранада инвагинация немесе төмпешік пайда болады. Ол қолбаға ұқсас, эноцитозды везикула деп аталады. Везикула мойыны қосылып мембранадан үзіледі. Везикула келешекте Гольджи комплексіне немесе лизосомаларға қарай тасымалдануы мүмкін, олармен қосылып, екінші лизосомалар немесе фаголизосомалар пайда болады.
1. Клетка үстіндегі заттар – антигендер
2. Клеткадан тыс матрикстегі заттар
3. Сигналды молекулалар (гормондар және медиаторлар)
Сыртқы және ішкі орта факторларына байланысты биомембрана қасиетінің өзгеруі мүмкін. Биомембрана қызметінің бұзылуы патологиялық процесс деп аталады, яғни биомембрана патологиясы. Биомембрана патологиясы мембраналық липидтермен, мембраналық белоктармен, рецепторлармен және гликогаликспен байланысады. Мысалы: улар, токсиндер, радиобелсенді және ультра күлгін сәулесі әсер еткенде биоммбрана нысана болып табылады. Мембраналық рецепторлар функциясының бұзылуы иммунды жүйенің патологиясына алып келеді. Мысалы: гипертония және атеросклероз. Бұл аурулардың түзілуі мембрана құрылымының бұзылысы, яғни белоктар және липидтердің қызметінің бұзылуымен тығыз байланысты. Қалыпты кездегі биомембрананың қызметін, қасиетін және құрылысын оқу, бізге оның негативті факторлардың әсері кезінде бұзылу механизмі себебі тереңірек түсінуге мүмкіндік береді.
Қолданылған әдебиеттер:
Негізгі
1. Стамбеков С.Ж., Петухов В.Л. Молекулалық биология. Оқулық/ҚР. Новосибирск: Семей МУ, 2003. –216 бет.
2. Әбилаев С.А. Молекулалық биология және генетика. Шымкент.2008, 424 б
3. Мушкамбаров Н.Н., Кузнецов С.Н. Молекулярная биология. Учебное пособие для студентов медицинских вузов, Москва: Наука, 2003,544 с.
4. Фаллер Д.М., Шилдс Д. Молекулярная биология клетки, Руководство для врачей. Пер с англ. М.: БИНОМ – Пресс,2003- 272 с.
5. Гинтер Е.К. Медицинская генетика. М., Медицина,2003.
6. Генетика. Учебник для ВУЗов / Под ред. Академика РАМН В.И. Иванова. – М.: ИКЦ «Академкнига»,2006.-638 с.:ил.
Қосымша:
1. Уилсон Дж., Хант Т. Молекулярная биология клетки. Сборник задач. Пер. с англ. –М.,Мир, 1994 -520 с.
2. Казымет П.К., Мироедова Э.П. Биология. Учебное пособие для студентов медицинских вузов. – Астана,2006,2007.
3. Медицинская биология и генетика/ Под.редакцией Куандыкова Е.У., Алматы,2004
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|