- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Внимание. Восприятие
ГЛАВА 20
Физиологические основы познавательной деятельности человека
Познавательной деятельностью человека называется получение и сохранение знаний. Эта деятельность осуществляется на основе базовых познавательных процессов, к которым относят внимание, восприятие, память, научение и мышление. Основным результатом познавательной деятельности человека является знание. В наиболее общей форме знание определяется как объекты, концепции и взаимосвязи, которые находятся в поле интереса (деятельности) человека. Знание человека имеет многочисленные проявления: умение, отношение к чему-либо или к кому-либо, информация, словарный запас, лексические правила, практические навыки и др.
Физиологической основой познавательной деятельности человека являются функции ассоциативных областей коры головного мозга. Эти области коры занимают большую ее площадь и имеют сложную синаптическую многоуровневую иерархию ядер (рис. 20.1).Иерархия ядер ассоциативных отделов коры включает в себя первичные сенсорные проекционные поля, вторичные унимодальные (имеют отношение к определенному виду чувствительности), гетеромодальные поля коры. В гетеромодальных полях многочисленные ядра, входящие в их состав, функционируют на основе разных видов информации. Уникальной функцией этих полей является связывание многочисленных унимодальных и трансмодальных полей в распределенные и интегрированные мультимодальные репрезентации, на основе ко-
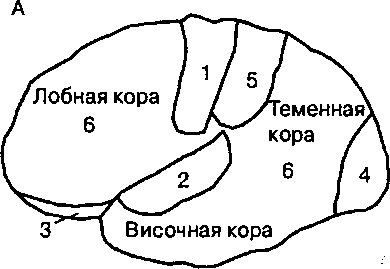
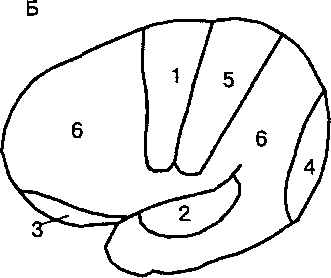
| Рис. 20.1. Схема расположения первичных проекционных и ассоциативных полей коры головного мозга человека (А) и шимпанзе (Б). У человека по сравнению с высшими приматами преимущественное развитие получили ассоциативные области коры, при участии которых осуществляются базовые познавательные процессы восприятия, научения и мышления. |
Первичные проекционные области коры: 1 — двигательная, 2— слуховая, 3 — обонятельная, 4 — зрительная, 5 — соматосенсорная, 6 — ассоциативная.
торого в мозге происходит превращение ощущения в познавательный процесс. Под репрезентацией (синоним — представление) понимают активацию нейронной сети ядер как унимодальных, так и гетеромодальных полей под влиянием поступающих в них сигналов. В познавательной деятельности человека функцией унимодальных ассоциативных полей коры головного мозга является восприятие базовых особенностей ощущений цвета, движения, формы и пространственной локализации стимулов внешней среды. Более сложное восприятие таких сенсорных признаков, как объекты, лица, словесные формы, пространственные локализации стимулов и последовательности звуков, осуществляется на более высоких уровнях синаптического переключения (4-й и 5-й уровни) унимодальных полей. Высшие синаптические уровни обработки сенсорной информации при познавательной деятельности происходят в гетеромодальных ассоциативных полях коры головного мозга, а также в паралимбической и лимбической коре. Эти поля коры головного мозга называются также трансмодальными. Трансмодальные поля коры головного мозга (средние отделы височной коры, зона Вернике, гиппокампально-энториальный комплекс, задняя теменная кора) выполняют в познавательной деятельности функцию трансформации: восприятия — в распознавание, словесных форм — в мысли, сцен и событий — в индивидуальный жизненный опыт человека.
Префронтальная ассоциативная кора в познавательной деятельности выполняет высшие функции мозга: вероятностное прогнозирование и программирование познавательной деятельности, анализ поступающей в этот отдел коры больших полушарий информации с учетом выполняемых результатов деятельности. Функции префронтальной коры в познавательной деятельности человека осуществляются на основе эксплицитной рабочей памяти. Механизмы этого вида памяти, для которой необходима точная и ясно выраженная или четко наблюдаемая человеком «информация», позволяют структурам префронтальной коры неоднократно обрабатывать в течение короткого времени одну и ту же осознаваемую человеком информацию и на этой основе включать ее элементы в содержание знания.
Познавательная деятельность человека обусловлена его сознанием. Сознание человека представляет собой психический процесс. Тем не менее сознание человека коррелирует с проявлениями активности (электрической или метаболической) в различных расположенных на расстоянии друг от друга ассоциативных полях коры головного мозга при базовых познавательных процессах внимания, восприятия, памяти и мышления.
20.1. Внимание
Сенсорные системы мозга человека одновременно воспринимают большое число влияний внешней среды. Одни из них имеют отношение к поведению человека в текущий момент времени, а другие — не влияют на таковое. Мозг человека не может воспринимать одновременно все многообразие воздействий внешней среды на организм. Поэтому в ЦНС происходят процессы, которые вычленяют лишь некоторые из стимулов внешней среды и «фильтруют» их от многих других. Это связано с тем, что организм человека способен эффективно функционировать во внешней среде, если происходит отбор или селекция тех стимулов, которые необходимы для осуществления его приспособления. Процессы отбора информации в мозге человека обозначаются понятием «внимание». Избирательная или селективная реакция организма человека на определенный вид сенсорной информации внешней среды называется вниманием. Внимание человека обусловлено функциями слуховой, соматовисцеральной, обонятельной, вкусовой и вестибулярной сенсорных систем. Внимание человека ответственно за выбор во внешней среде объектов и их локализации, о которых затем необходимо получить более подробную информацию и, одновременно, исключить из восприятия объекты другой локализации. Внимание человека одновременно не способно ориентироваться на несколько стимулов внешней среды или несколько мыслительных процессов.
20.1.1. Формы внимания
Избирательный отбор одного стимула из множества раздражений внешней среды происходит у человека на основе непроизвольного (пассивного, или рефлекторного) и активного, или произвольного, внимания.
Непроизвольное внимание представляет собой автоматическую, рефлекторную реакцию организма человека (как правило, поворот глаз и/или головы в направлении источника раздражения) в ответ на действие стимулов внешней среды. При этом стимулы внешней среды, которые способны вызвать внимание человека, должны быть очень сильными либо значительными, либо резкими и неожиданными, и иметь конкретную природу — световые, звуковые, обонятельные или ноцицептивные (например, внешняя или внутренняя боль). Непроизвольное внимание возникает неосознанно и не требует умственных усилий со стороны человека для направления взгляда или поворота головы в сторону источника раздражения. Как правило, непроизвольное внимание обусловлено такими качествами стимулов, как интенсивность или неожиданность их возникновения для человека, или их новизна.
Произвольное внимание у человека представляет осознаваемое человеком ориентирование органов чувств, как правило, органа зрения, на восприятие конкретного объекта или явления внешней среды. Основным видом произвольного внимания является зрительное произвольное внимание, которое подразделяется на пространственное и объект-ориентированное. Пространственное зрительное внимание представляет собой процесс изменения зрительного внимания с одного объекта на другой. Направленность зрительного внимания на конкретный объект называется объект-ориенти- рованным зрительным произвольным вниманием (синоним — фокусное внимание).
Произвольное внимание возникает в том случае, если человек осознает действие (поворот глаз и/или головы), направленное на рассматривание, выслушивание или осязание какого-то объекта или явления внешней сре-
Таблица 20.1. Две формы зрительного внимания человека
| Свойства внимания | Формы внимания | |
| непроцзволное | произвольное | |
| Пространственная специфичность Детальная специфичность Продолжительность Произвольный контроль Зависимость от задания | Через поле зрения Действует в любое время и во всех пространственных направлениях Временное Отсутствует Не зависит | Пространственно обусловленное (фокусное) Может избирать специфичный атрибут Устойчивое (с произвольным усилием) Обязателен Зависит |
ды. Произвольное внимание у человека обусловлено предшествующим научением (ассоциативное научение, инструкция, задание, внутреннее желание). Произвольное внимание человек может направлять на мыслительные процессы, обусловливающие, например, принятие решения и последующие за этим действия. По сравнению с произвольным вниманием, непроизвольное внимание возникает независимо от инструкций или внутреннего намерения человека (табл. 20.1).
20.1.2. Нейрофизиологические механизмы внимания
Физиологическая основа внимания состоит в том, что оно обусловлено увеличением и синхронизацией нейронной активности в тех отделах ЦНС, которые воспринимают сенсорные стимулы, и увеличением активности структур теменной и лобной коры мозга человека.
Наиболее полное проявление избирательности, или селективности, внимания связано с функцией зрения у человека. Зрительное пространство содержит множество стимулов, специфическая локализация которых является избирательной для процесса зрительного внимания. Емкость зрительного внимания ограничена. Своеобразной ценой избирательности внимания является «слепота» к зрительным стимулам, на которые человек не обращает внимания. При этом зрительное внимание способно выбирать наиболее интересный объект в поле зрения или изменять «фокус» внимания с одного объекта на новый. Наиболее важной функцией непроизвольного и произвольного зрительного внимания является быстрое направление взгляда человека на интересующий объект в окружающем зрительном пространстве. Физиологические механизмы непроизвольного и произвольного внимания человека тесно взаимосвязаны между собой. У человека нейронные структуры внимания представлены отделами верхних бугров четверохолмия, подушки таламуса, зрительной коры, лобного глазного поля, дополнительного глазного поля лобной коры, задней теменной коры, нижней височной и префронтальной коры. Все отделы зрительных областей коры головного мозга играют важную роль в зрительном внимании. Однако наибольший дефицит зрительного внимания у человека возникает при повреждениях теменных долей коры. Это проявляется в игнорировании контралатерального зрительного пространства, которое возникает чаще при повреждении правой теменной области коры головного мозга и реже — левой. Поэтому правая теменная область коры головного мозга выполняет доминирующую функцию во внимании человека. Физиологические механизмы внимания обусловлены функцией следующих подкорковых и корковых центров внимания.
20.1.2.1.Функции среднего мозга и моста в контроле внимания
Функция состоит в контроле движения глаз и/или головы человека в направлении источника раздражения. Этот механизм лежит в основе обеих форм внимания человека, т. е. непроизвольного (ориентировочный рефлекс по И. П. Павлову) и произвольного.
Ориентированное движение глаз при внимании осуществляется синергичными сокращениями трех пар глазодвигательных мышц. Нейроны, которые иннервируют мышечные волокна этих мышц, образуют III (глазодвигательный), IV (блоковой) и VI (отводящий) черепные нервы (рис. 20.2).
В стволе мозга, на уровне моста, расположена нейронная сеть ретику-
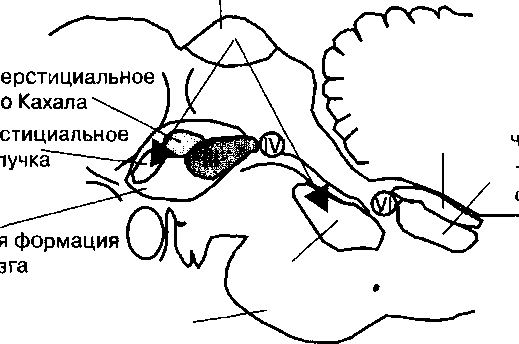
| Верхние бугры четверохолмия |
| Ростральное интер ядро продольного г |
| И нт ядр |
| Мост |
| Рис. 20.2. Нейронный контроль саккадических движений глаз на уровне структур среднего мозга. Направление движения глаз (саккад) управляется мотонейронами двух центров взгляда ретикулярной формации. В парамедианной ретикулярной формации моста расположен центр горизонтального направления движения вгляда, а в ростральном интерстициальном ядре ретикулярной формации находится центр вертикального направления движения взгляда. Ill, IV, VI — ядра глазодвигательного, блокового и отводящего нервов, которые иннервируют мышцы глазных яблок человека. Стрелками показано направление регулирующих влияний структур верхних бугров четверохолмия. |
| Мозжечок |
| Таламус |
| Ретикулярна! среднего мо: |
| Ядро подъязычного нерва |
| Ретикулярная формация продолговатого мозга |
| Парамедианная ретикулярная формация моста |
лярной формации, функция которой состоит в регуляции многих аспектов внимания: генерация быстрых (саккадических) движений глаз (парамедианная ретикулярная формация); участие в произвольной регуляции направления взгляда человека (ретикулярные нейроны вблизи ядра отводящего нерва); генерация горизонтального направления движения взгляда (нейроны ретикулярной формации моста и прилегающие к ядру подъязычного нерва); содружественное горизонтальное направление движения взгляда обоих глаз. Последний комплекс ядер в соответствии с его функцией получил название центр горизонтального направления движения взгляда человека. Аналогичная нейронная сеть, или центр вертикального направления движения взгляда, расположен в области интерстициального ядра Кахала.
Центры горизонтального и вертикального направления движения взгляда человека регулируют поворот глазных яблок через мотонейроны ядер III, IV и VI черепных нервов и иннервируемые ими мышцы.
Функция верхних бугров четверохолмия. Верхние бугры четверохолмия представляют собой область ствола мозга, которая образует основный выход к ретикулярным нейронным сетям моста и среднего мозга, при участии которых возникает движение или остановка взгляда человека при внимании.
Зрительные сигналы от сетчатки гдаз поступают к нейронам верхних слоев верхних бугров четверохолмия. Аксоны этих нейронов направляются к нейронам глубоких слоев верхних бугров четверохолмия, а также к таламусу и стволовым ядрам. Функция этих «нисходящих» связей заключается в том, что нейроны глубоких слоев верхних бугров четверохолмия становятся высокоактивными перед началом движения глаз и головы в направлении источника раздражения во внешней среде. Высокочастотная активность нейронов глубоких слоев верхних бугров четверохолмия резко активирует глазодвигательный механизм человека через нейроны ретикулярных
ядер ствола мозга при возникновении реакции внимания (поворот глаз и/ или головы человека). Поэтому глаза человека способны в течение нескольких десятков миллисекунд ориентироваться на объект после раздражения рецепторов сетчатки.
Функция нейронов глубоких слоев верхних бугров четверохолмия является мультимодальной при внимании, поскольку эти отделы получают сигналы от зрительной коры, от слуховых, соматосенсорных релейных ядер и других ядер ствола мозга. Глубокие слои нейронов верхних бугров четверохолмия имеют строгую топографическую организацию в виде нейронных карт. Это проявляется в том, что при возбуждении нейронов определенной локализации участков глубоких слоев верхних бугров четверохолмия возникают строго координированные движения глаз и поворот головы.
20,1.2.2, Функции корковых центров внимания
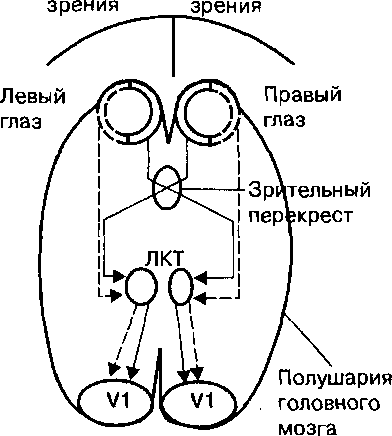
Простое произвольное изменение внимания с одной точки зрительного пространства на другую с помощью движений глаз активирует у человека большую нейронную сеть коры головного мозга, которая включает комплекс зрительная кора — средняя теменная область, лобные глазные поля, дополнительные глазные поля, часть передней поясной извилины, заднюю затылочную кору (рис. 20.3).Функции этой обширной нейронной сети при внимании заключаются в контроле движения глаз и головы человека через верхние бугры четверохолмия. Структуры верхних бугров четверохолмия и корковых центров внимания функционируют параллельно. В обычных физиологических условиях возникновение активности в нейронах верхних бугров четверохолмия находится под контролем лобных зрительных полей, задней теменной коры и нейронов ретикулярной части черной субстанции ствола мозга.
Функции комплекса структур стриарной зрительной коры и средней теменной области (зона МТ) в механизмах внимания состоят в проведении зрительной информации о направлении и скорости движения стимулов в зрительном поле к структурам задней теменной коры головного мозга. Из структур экстрастриарной коры информация передается в лобную кору, а именно в лобные и дополнительные глазные поля, которые выполняют основную функцию в планировании и выполнении ориентированных движений глаз. Структуры первичной зрительной коры выполняют основную функцию в объект-ориентированном внимании человека и в выборе или селекции объекта глазодвигательной системой мозга человека с того момента, как только цель оказалась в сенсорном поле. В ответ на зрительный стимул стриарная затылочная кора человека активируется довольно быстро, примерно через 40 мс.
Роль теменной коры человека в механизмах внимания. При внимании теменные отделы мозга человека, а именно структуры внутритеменной борозды и нижней теменной доли, повышают уровень метаболической активности. Эта область коры головного мозга является центром, который вовлекается в поддержание внимания специфической пространственной локализации или, как правило, в генерацию сигналов в глазодвигательной системе мозга при изменении внимания с одной локализации на другую. Эту функцию отделы задней теменной коры выполняют через структуры верхних бугров четверохолмия.
Нейроны задней теменной коры реагируют строго топографически на зрительные стимулы и участвуют в произвольном изменении пространст-
Произвольное зрительное внимание (произвольное движение глаз)
Непроизвольное зрительное внимание
Пространственное зрительное внимание
| зтк |
| кора |
| Мозжечок |
| Зрительная информация |
| Двигательные программы |
| Генератор саккад |
| Интеграция пространственной зрительной информации |
| Ретикулярная формация |
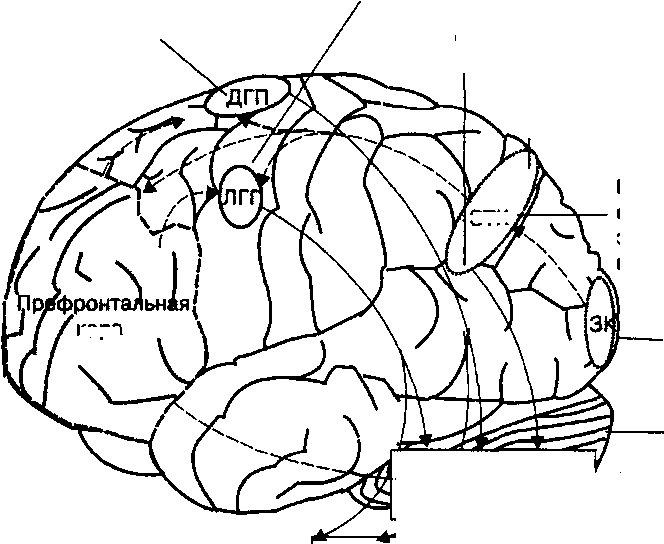
| Рис. 20.3. Функции высших центров внимания, расположенных в коре головного мозга человека. |
| Верхние бугры четверохолмия |
Центры внимания коры головного мозга человека включают зрительную кору (ЗК), которая направляет зрительную информацию в заднюю теменную кору (ЗТК). Этот корковый центр внимания регулирует механизм непроизвольного зрительного внимания, осуществляет интеграцию пространственной зрительной информации. Задняя теменная кора (преимущественно правая) направляет сигналы в лобное глазное поле (ЛГП) и дополнительное глазное поле (ДГП). ЛГП функционирует как центр произвольного зрительного внимания человека и выполняет двигательные программы при участии ДГП. Префронтальная кора включена в произвольную регуляцию внимания путем коррекции функции центров лобного и дополнительного зрительного полей. Человек при участии структур префронтальной коры способен тормозить непроизвольное внимание. Верхние бугры четверохолмия являются основным подкорковым центром регуляции непроизвольного (рефлекторного) и произвольного внимания при участии различных сенсорных систем и под влиянием двигательных команд центров внимания коры головного мозга.
венного внимания. Это проявляется прежде всего в том, что при изменении направления взгляда на новое положение рецептивные поля нейронов задней теменной коры также перемещаются на новое положение. Более того, рецептивные поля задней теменной коры способны занимать новое положение до начала движения глаз и изменения направления взгляда на новый объект, а следовательно, нейроны этого коркового центра внимания получают сигналы из других отделов коры относительно предполагаемого движения глаз в новом направлении. Поэтому активность нейронов задней теменной коры как основного высшего центра внимания всегда нарастает при ориентировании взгляда на объекте, если человек ожидает появления объекта в зрительном поле или думает о появлении в нем такого объекта. После начала действия зрительного стимула зрительная информация достигает теменных отделов коры примерно через 60 мс и передается к этим структурам от стриарной коры. При этом активация структур стриарной и
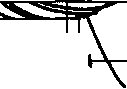
Рис. 20.4. Функция ретикулярной части черной субстанции и базальных ганглиев в контроле внимания.
Ретикулярная часть черной субстанции (ЧС) ствола мозга получает вход от хвостатого ядра (ХЯ) базальных ганглиев и функционирует как «ворота» при произвольном контроле движения глаз. При отсутствии зрительных стимулов в поле зрения ЧС тонически тормозит структуры верхних бугров четверохолмия, что препятствует возникновению нежелаемых движений глаз и изменению направления взгляда. Непосредственно перед началом произвольного изменения взгляда в черную субстанцию из базальных ганглиев поступают импульсы, которые снижают торможение верхних бугров четверохолмия. Активация базальных ганглиев происходит под влиянием двигательных команд из центров внимания коры головного мозга. В результате становится возможным произвольное изменение направления взгляда. Знак плюс обозначают активацию центров регуляции направления взгляда со стороны верхних бугров четверохолмия и торможение, а знак минус — торможение верхних бугров четверохолмия со стороны ЧС.
| Центры горизонтального и вертикального направления движения взгляда |
Функция лобных глазных полей заключается в планировании движения глаз при изменении направления взгляда. Лобные глазные поля расположены в прецентральной борозде лобных долей мозга человека. Реакции нейронов этой области коры головного мозга организованы топографически: нейроны активируются только в том случае, если объект находится в определенной части зрительного поля. Функция лобных глазных полей осуществляется при помощи проекций аксонов нейронов этого отдела коры к структурам верхних бугров четверохолмия и стволовым премоторным (ретикулярным) нейронам. Лобные зрительные поля являются корковым премоторным центром в регуляции внимания и обеспечивают выбор целей для движений глаз и изменения направления взгляда среди многих зрительных целей в окружающем человека пространстве. Функцией этой области коры головного мозга является торможение непроизвольных движений глаз, а также генерация произвольных, не связанных со зрительными стимулами движений глаз (например, при мышлении).
Функцией дополнительных глазных полей коры является выполнение движения глаз в направлении объекта внимания. Дополнительные глазные поля расположены медиальнее лобных глазных полей, вдоль межполушарной борозды на дорсомедиальной поверхности лобных долей. Нейроны этого коркового центра внимания обладают пространственной чувствительностью и регулируют движение глаз таким образом, чтобы объект или его какая-то часть находились в центре поля зрения. Благодаря функции структур дополнительного глазного поля у человека осуществляется объект-ори- ентированное внимание.
Функция базальных ганглиев. В механизме произвольного внимания функция базальных ганглиев состоит в активации структур ретикулярной части черной субстанции среднего мозга (рис. 20.4).Нейроны ретикулярной части черной субстанции функционируют подобно «воротам» относительно функции структур верхних бугров четверохолмия при внимании. Так, при отсутствии в окружающем пространстве вызывающих внимание зрительных стимулов нейроны черной субстанции тормозят нейроны верхних бугров четверохолмия и препятствуют, таким образом, случайному изменению направления взгляда. Под влиянием произвольных двигательных команд из лобной коры, поступающих к нейронам черной субстанции через структуры хвостатого ядра базальных ганглиев, тоническое торможение нейронов верхних бугров четверохолмия снижается, и они приобретают способность регулировать произвольные движения глаз в направлении объекта в поле зрения.
Дорсолатеральные области префронтальной коры обеспечивают произвольный контроль различных форм поведения, в том числе ориентированные движения глаз и головы на интересующий человека объект в окружающем пространстве. При участии префронтальной коры осуществляется произвольная задержка внимания, в результате которой человек осознанно не производит объект-ориентированное движение глаз и головы в направлении источника сильного сенсорного раздражения.
20.1.3. Внимание при различных модальностях
Внимание у человека проявляется в отношении стимулов разной модальности, т. е. зрительных, слуховых, соматосенсорных, обонятельных и вестибулярных. Слуховые центры ствола мозга взаимодействуют с системой регуляции зрительного внимания на уровне структур верхних бугров четверохолмия (ориентировочный рефлекс), а на уровне коры — с теменными областями коры, преимущественно со структурами нижней теменной доли в правом полушарии мозга. Активация структур этой области коры головного мозга происходит при бинауральном и дихотическом звуковом воздействиях. Соматосенсорная система на подкорковом уровне (через верхние бугры четверохолмия) и на уровне коры при участии структур соматосенсорной коры также взаимодействуют с системой регуляции внимания через отделы задней теменной коры.
Наибольшее внимание вызывает стимул одной модальности (зрительной, слуховой или соматосенсорной), т. е. внимание эффективнее осуществляется при участии конкретной сенсорной системы. Однако, несмотря на наличие внимания к различным внешним стимулам, в коре головного мозга человека имеется одна единая надмодальная (или мультимодальная) система регуляции внимания. Основным мультимодальным центром внимания является задняя теменная кора головного мозга. Функция задней теменной коры при внимании проявляется в том, что пространственное слуховое или пространственное зрительное внимание у человека всегда направлены на один и тот же объект в окружающем пространстве. При произвольном изменении пространственного слухового внимания в направлении наилучшего восприятия звукового стимула в эту же область направляется пространственное зрительное внимание, и наоборот. При участии задней теменной коры каждая сенсорная система вызывает эквивалентное изменение пространственного внимания у человека, что способствует увеличению процессов локализации внимания на определенной цели в условиях активации различных сенсорных систем мозга.
20.2. Восприятие
Восприятие — это процесс трансформирования физического раздражения в физиологическую информацию. Процессы восприятия сенсорной информации различной модальности осуществляется в первичных и вторичных проекционных областях коры головного мозга. Наиболее изученными видами восприятия у человека являются зрительное и слуховое, в меньшей степени — соматосенсорное. Механизмы восприятия запаха и вкуса изложены в главе 17 «Сенсорные системы».
20.2.1. Зрительное восприятие
Человек распознает новый объект, появившийся в поле зрения, в течение долей секунды. При этом зрительное восприятие включает оценку контраста, лица, объектов, зрительного пространства, а также единого изображения, воспринимаемого обоими глазами (бинокулярное зрение).
Структуры ЦНС, которые участвуют в «зрительном поведении» человека, широко представлены в коре головного мозга, однако не все из них вовлечены в процесс восприятия. Например, двустороннее повреждение височных долей коры вызывает выраженные нарушения в зрительном распознавании и памяти, но при этом остается неизмененным зрительное восприятие. Напротив, повреждение у человека затылочных долей вызывает слепоту. Небольшой латентный период и длительность реакций нейронов первичной зрительной коры (порядка 40 мс) на зрительные стимулы свидетельствуют об основной функции этого отдела зрительной коры в восприятии зрительной информации.
20.2.1. L Функции стриарной коры в зрительном восприятии
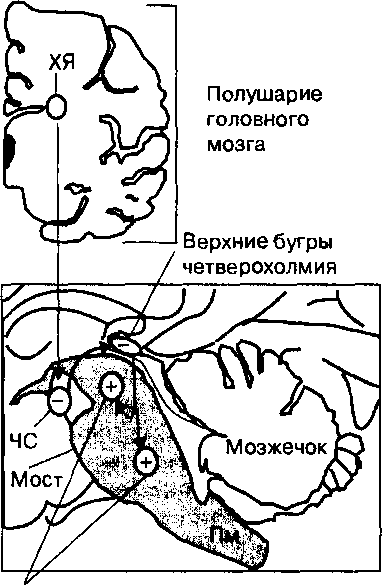
Первичная зрительная кора (стриарная кора, поле VI) является первым корковым уровнем, на котором конвергируют сигналы от сетчатой оболочки обоих глаз человека, и последним полем в зрительной системе, нейроны которого реагируют непосредственно на зрительные стимулы (рис. 20.5).Первичная зрительная кора представляет собой единственную структуру коры головного мозга человека, которая получает сигналы непосредственно от нейронов латеральных коленчатых
| Левое поле Правое поле |
Рис. 20.5. Схема передачи импульсов от сетчатки к нейронам зрительной коры (поле VI).
Пути от латеральных коленчатых тел (Л КТ) в правую и левую области зрительной коры состоят из относительно равных проекций от сетчатки обоих глаз. Нейроны первичной зрительной коры активируются на зрительные стимулы и обеспечивают функцию зрительного восприятия.
(глубины), ориентационных стимулов, контуров объектов, движения, формируется глазодоминантность. Нейроны стриарной коры имеют высокую чувствительность к световому контрасту, а их реакции прямо взаимосвязаны с порогами светового контраста (описаны в главе 17 «Сенсорные системы»). Восприятие освещенности предмета также является функцией стриарной коры. Структуры полей V1/V2 являются высшими зрительными центрами восприятия, в которых начинается восприятие бинокулярного зрения, хотя в этих центрах зрительного восприятия расположено меньше нейронов с подобными свойства, чем в последующих зрительных полях. Восприятие стереоопсиса происходит в результате взаимодействия на нейронах стриарной коры монокулярной зрительной информации, возникающей в результате перекрытия одного объекта другим в поле зрения.
К своеобразной форме зрительного восприятия относится воображение человека в отсутствие какой-либо световой стимуляции сетчатки глаз. При этом у человека активируются ретинотопические поля зрительной коры, а не ее ассоциативные поля.
20.2.1.2. Зрительное восприятие при участии отделов экстрастриарной коры
Нейроны зрительного поля VI моносинаптически связаны с нейронами других ретинотопических зрительных областей коры — V2, V3/VP, V3A, V4v и МТ фис. 20.6).
При произвольном или непроизвольном внимании активируются высшие зрительные корковые поля, преимущественно поля V2 и V4.
Двигательная кора
| Лобная кора |
| V2 |
| МТ |
| Височная кора |
| Боль |
| Первичная зрительная кора (поле V1) |
| VP |
V7 V3A
V8 V4v Восприятие цвета Восприятие лица Запах Слуховая кора
Рис. 20.6. Схематическое изображение локализации функций познавательной деятельности человека в коре головного мозга: восприятие (по данным позитронноэмиссионной томографии и функционального магнитного резонансного исследования).
На рисунке отмечены участки коры головного мозга человека, в которых возрастает локально мозговой кровоток при выполнении человеком заданий на восприятие. VI, V2, V3, V3A, V/P, V4v — классические ретинотопические поля зрительной коры человека, функцией которых является восприятие относительно простых зрительных стимулов, поля V7 и V8 — имеют слабо выраженную ретинотопическую организацию. За пределами этих полей расположены области зрительной коры, не имеющие ретинотопической организации и поэтому выполняющие «глобальные» функции восприятия движения, пространства, цвета и формы объектов внешней среды. МТ — средняя височная область.
После восприятия зрительного раздражения структурами поля VI, далее, в каждом последующем ретинотопическом поле, происходит копирование 'Д карты поля VI. Поэтому при повреждении полей V2, V3/VP и V4v у пациентов возникает выпадение восприятия */4 части зрительного поля (квад- ранопсия), а поля VI и V3A —половины зрительного поля (гемианопсия). Структуры полей V2, V3/VP, V3A, V4v и МТ зрительной коры образуют второй синаптической уровень и относятся к восходящим ассоциативным зрительным полям, функцией которых является распознавание относительно простых параметров зрительных сигналов. Кпереди от промежуточных полей зрительной коры (поля V7 и V8), которые имеют слабо выраженную ретинотопическую карту, расположены структуры зрительной коры, которые не имеют ретинотопической организации и поэтому выполняют «глобальные» функции зрительного восприятия движения, цвета, лиц, участков зрительного поля и кинетику движения граничащих между собой элементов.
В процессе зрительного восприятия у человека функции этих отделов зрительной коры количественно разделены на теменные и височные информационные процессы («потоки»). Структуры средней теменной области (МТ), верхней теменной области, полей V3A и V7 формируют у человека теменной (дорсальный) путь или поток, который обеспечивает восприятие движения и пространства.
Вентральный (височный) «поток» информации (структуры полей V8, лицевого поля веретенообразной извилины) выполняют функцию восприятия более сложно организованных особенностей зрительной информации, таких как цвет и форма объектов.
20.2.1.3. Особенности зрительного восприятия индивидуальных лиц и объектов Зрительное поле, специализирующееся на восприятии лица, расположено в пределах средней части веретенообразной извилины (поля 37 и 20 по Брод- ману). Нейроны веретенообразной извилины активируются в большей степени при рассматривании анатомически нормальных и вертикально расположенных лиц, чем перевернутых или хаотично расположенных в поле зрения объектов. В то же время степень активации «лицевого» поля коры не различается при рассматривании знакомых и незнакомых лиц. Нейроны «лицевого» поля распознают лица на категориальном или генетическом уровне, т. е. до стадии индивидуального распознавания лица.
Непосредственно латеральнее веретенообразной извилины расположено поле, нейроны которого специализируются на распознавании словесных форм и слов, связанных в последовательный ряд при их написании. У человека письменный язык является поздним филогенетическим приобретением, и в онтогене
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|