- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Методы определения морфометрических параметров хвои
2. Методы определения морфометрических параметров хвои
К числу важнейших признаков древесных растений относят характеристики листового аппарата. Их связывают с эффективностью фотосинтеза, продуктивностью и устойчивостью растений к лимитирующим экологическим факторам (Барская, 1967; Озолина, 1972; Лир, 1974; Либберт, 1976; Нечипорович, 1982; Крамер, 1983; Ермолова, 1998; Duysens, 1961; 1963; Lichtentaller, 1983; Naidu, 1993; Niinemets, 1997; 2002 a, b; Gordon, 1998; Porté, 1998; Kayama, 2002; Poljanšek, 2011), в том числе у сосны обыкновенной (Правдин, 1964; Rosemann, 1991; Wingsle, 1991; Jach, 2000; Vanninen, 2000; Niinemets, 2001; 2002 b; Lamppu, 2002; Taulavuori, 2010). Не менее важны эти показатели в решении идентификационных и классификационных задач, притом, что характеристики разновозрастной хвои не одинаковы (Niinemets, 1997; Porté, 1998; Kayama, 2002; Liber, 2002).
Морфология хвои представляет собой предмет многоплановых исследований важнейших лесообразующих пород. Отмечена корреляционная зависимость степени развития хвои и темпов роста растений в насаждениях (Луганская, 1976), а её варьирование в высокопродуктивных насаждениях по ряду признаков отнесена к очень высокому уровню изменчивости (Мамаев, 1969 а, б; Лебков, 2004 а). Наиболее стабильным из количественных признаков хвои является её длина (Мамаев, 1969 а), по другим сведениям длина, ширина и их соотношение, а также ширина клеток эпидермиса (Boratyńska, 2000; Klõšeiko, 2008). Вместе с тем Л.Ф. Правдин (1972) отмечал заметную зависимость этого показателя от внешних условий при выраженном контроле со стороны генотипа растений. Под высокой степенью генетического контроля и окружающей среды (без особого взаимодействия генотип – окружающая среда) находятся скорость роста и длина хвои (Райт, 1978). У северных форм хвоя короче, чем у южных (Царев, 2003). Ряд авторов указывает на низкий уровень изменчивости линейных показателей хвои, в том числе внутриклоновой (Чернодубова, 1996; Ткаченко, 2000). Другие исследователи, напротив, отмечают значительный уровень изменчивости линейных параметров хвои в популяции под действием экологических факторов (Шульга, 1970; Lamppu, 2002). Считают, что размер хвои является косвенным показателем энергии роста сосны в географических культурах Казахстана: климатипы (климатического экотипа) с более длинной хвоей и повышенным количеством боковых почек на осевом побеге образуют более мощную крону (Бреусова, 1970). Кроме того, известно, что показатель вариабелен в пределах отдельных особей климатипа, а также изменяется по годам (Бреусова, 1970). Аналогичные сведения приводятся по Бельгии (Cermák, 1998), Финляндии (Salminen, 2006). Не установлено изменение длины хвои в естественных популяциях в зависимости от географической широты (Пугач, 1975), притом что представители разных популяций неоднородны в этом отношении (Urbaniak, 2002 a, b; 2003). Вместе с тем сообщается о географической внутривидовой изменчивости европейских популяций по содержанию питательных веществ и микроэлементов в хвое (Oleksyn, 2002; Sariyildiz, 2006), общему содержанию азота, морозостойкости и оксидантному стрессу (Taulavuori, 2010), о возможности использования морфологических параметров (Androsiuk, 2011) и анатомического строения хвои (Urbaniak, 2008) в качестве индикаторов географического происхождения популяций (Urbaniak, 2008; Androsiuk, 2011), как для сосны обыкновенной, так и для других видов этого рода (Boratyńska, 2004). Внутрипопуляционная изменчивость морфологических и анатомических признаков хвои сосны обыкновенной низкая (Boratyńska, 2000; 2003).
Длина хвои коррелирует с темпами роста: чем длиннее хвоя, тем выше темпы роста (Мольченко, 1982). С темпами роста связывают и форму хвои: у быстрорастущих она скрученная (Альбенский, 1959). Масса хвои у всех клонов варьирует значительно (Cv = 12…30 % и более), что может служить основанием для отбора потомства по этому признаку; разработана шкала изменчивости по длине и массе (Ткаченко, 2000; Лебков, 2004а). Отмечена высокая степень наследуемости различий по массе сухой летней хвои (Райт, 1978). Мощность покровных тканей хвои зависит от географического происхождения и влияет на поражаемость болезнями (О.Г. Капер, цит. по Цареву, 2003). Содержание макроэлементов в хвое претерпевает изменение по ярусам кроны и в течение вегетационного периода (Кищенко, 2000; Терехова, 2002). Продолжительность жизни хвои сосны обыкновенной изменчива и может достигать 5 – 7 лет (Шульга, 1970; Туллус, 1991; Евдокимов, 2003). Л.Ф. Правдин (1972) предлагает в этой связи разделить ареал сосны на две части: западную с малой продолжительностью жизни хвои и восточную – с большой.
Представленный материал свидетельствует об отсутствии однозначных трактовок накопленной информации о свойствах хвои, позволяет признать необходимость продолжения исследований в отмеченном направлении. Их актуальность, помимо прочего, обусловлена ограниченностью сведений о подобных характеристиках плюсовых деревьев сосны обыкновенной на территории Российской Федерации.
Началу работы предшествует выдвижение гипотез:
1) клоновые репродукции плюсовых деревьев сосны обыкновенной существенно различаются по параметрам хвои как в свежесобранном, так и воздушно-сухом состоянии;
2) различия между плюсовыми деревьями обусловлены генотипически;
3) параметры хвои весьма чувствительны к влиянию факторов среды.
Исследования можно строить в апробированном нами порядке (Бессчетнов, 2012 б; Бессчетнова, 2004 е; 2004 ж; 2012 а; 2014 а). На каждом опытном участке каждое плюсовое дерево (ортет) было представлено постоянным числом учетных растений (рамет), количество которых по разным объектам ПЛСБ и ЕГСК составляло 3 – 9 шт. При этом обеспечивалась территориальная полнота и равномерность их размещения на площади анализируемых лесосеменных плантаций и архивов клонов. Количество ортетов определено проектной документацией и схемами создания обследованных объектов постоянной лесосеменной базы и единого генетико-селекционного комплекса. Хвоя заготавливалась по 50 – 100 пар с нескольких (не менее 3) разных побегов каждого учетного дерева. Учетные побеги (в количестве не менее 3) выделяли в среднем поясе периферии хорошо освещенного участка кроны (предварительно визуальным способом устанавливалось, какая сторона наиболее полно освещена в течение светлого времени суток). Из их центральной части одновременно и равномерно заготавливалась полностью вызревшая (на момент завершения вегетационного периода) нормально развитая одновозрастная хвоя (1-летняя и 2-летняя раздельно), не имевшая признаков повреждения. Она служила источником формирования первичных единиц выборки; количество, размер и структура выборок зависела от типа анализа. Период проведения работ – 2006 – 2014 гг.
Первичной единицей выборки служили: для определения длины свежесобранной и воздушно-сухой хвои – отдельная хвоинка; для определения массы – отдельный пучок из 2 хвоинок. При оценке длины хвои для разграничения каждой из двух хвоинок в их пучке и раздельной регистрации в полевых журналах им условно присваивался учетный индекс: «1-я хвоинка» или «2-я хвоинка». Их длину измеряли линейкой с точностью до 0,5 мм, массу определяли взвешиванием на торсионных весах ВТ-500 и трехразрядных электронных весах Acculab Vicon VIC-300d3 с точностью до 0,001 г. Повторный учет значений проводился после длительного (4 месяца) раздельного высушивания образцов в специально подготовленных контейнерах при постоянном свободном доступе воздуха в сухом помещении при комнатной температуре. Указанный режим был выбран исходя из представлений о возможном удалении из хвои части летучих веществ (например, смол и эфирных масел) под воздействием высоких температур режима принудительной термической сушки. Это могло бы отрицательно сказаться на точности проводимых исследований (на данном этапе мы фиксировали только потерю воды). После этого навески доводились до абсолютно-сухого состояния в сушильных шкафах при температуре 105оС в течение 8 часов (до достижения постоянной массы). Количество воды определяли как разность между результатами взвешиваний свежесобранной и сухой хвои (в воздушно сухом и абсолютно сухом состоянии), её долю устанавливали относительно исходной массы, также вычисляли отношение массы к длине хвои (распределенная масса).
Принятый порядок отбора первичных единиц выборки и формирования выборочных совокупностей позволял организовать одно-, двух- и трех факторные иерархические дисперсионные комплексы (рис. 2.1).

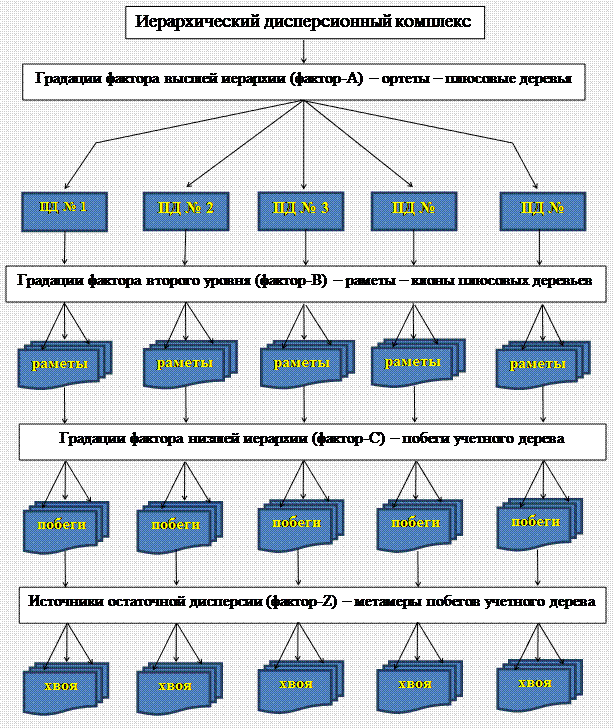
Рисунок 2.1 – Схема организации трехфакторного иерархического дисперсионного комплекса, формируемого в пределах
отдельного объекта ПЛСБ или ЕГСК
Уровнями иерархии в них (см. рис. 2.1) служили следующие источники варьирования: уровень высшей иерархии, формируемый фактором А – различия между собственно плюсовыми деревьями (ортетами); второй уровень иерархии, формируемый фактором В – различия между учетными деревьями-клонами (раметами); третий уровень иерархии, формируемый фактором С – различия между учетными побегами одного учетного дерева. Источником остаточной (случайной) дисперсии, формируемой фактором Z, являлись различия между первичными единицами выборки – метамерами или хвоей (Бессчетнова, 2012 а).
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|