- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Отступление №2. Отступление №3
Отступление №2
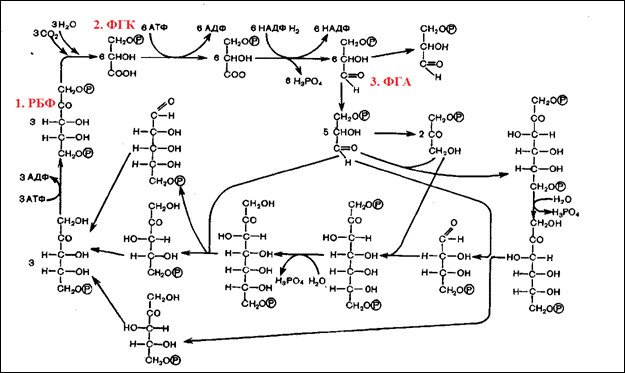 Рассмотрим C3 (так же Цикл Кальвина или Восстановительный пентозофосфатный цикл) путь (именно его изучают в школе. он самый распространённый):
Рассмотрим C3 (так же Цикл Кальвина или Восстановительный пентозофосфатный цикл) путь (именно его изучают в школе. он самый распространённый):
(начало реакций - 1 Рибулозо-1, 5-бисфосфат)
Протекает в строме хлоропласта.
1. В строме хлоропласта имеются молекулы рибулозо-1, 5-бисфосфата (1 на рисунке) и особый фермент RuBisCO. Этот фермент способен к акцепции (захвату) CO2 из окружающей среды (стромы). Именно это он и делает.
Отступление №3
Рибулозо-1, 5-бисфосфат имеет углеродный 5 атомный скелет и 2 фосфата на замыкающих атомах. RuBisCO" прицепляет" захваченный им CO2 к рибулозо-1, 5-бисфосфату, в результате чего образуется крайне нестабильное C6 соединение, которое быстро распадается на две молекулы фосфоглицериновой кислоты. (2)
2. Далее при взаимодействии с 2NADP . H2 и 2ATP из 2 молекул фосфоглицериновой кислоты выходит 2 молекулы фосфоглицеринового альдегида (3). Соответственно выходит ещё 2NADP+ и 2ADP, которые отправляются на дальнейшее использование (в световые реакции).
Фосфоглицериновый альдегид является полноценным углеводом. Автотроф (да, цикл Кальвина имеют и многие хемоавтотрофы) может отправить фосфоглицеральдегид на глюконеогенез (синтез глюкозы) или использовать сразу (например, провести клеточное дыхание, сделав из него сначала пируват).
Мы рассмотрели течение реакции для 1 молекулы рибулозо-1, 5-бисфосфата. Но как вы видите по схеме, в действительности полноценный цикл Кальвина требует 3 молекулы РБФ и даёт на выходе из нашего пункта 6 молекул ФГА. Это требуется для дальнейших реакций:
3. И так мы имеем 6 молекул ФГА. 1 из них можно забрать как раз для использования. Это и будет выгода автотрофа со всего фото(хемо)синтеза. А вот оставшиеся 5 ФГА идут в дальнейшие реакции цикла Кальвина (см. схему). Все их разбирать мы не будем. В этом нет необходимости для понимания фотосинтеза. Но пару слов скажем.
В результате всех дальнейших реакций из 5 молекул ФГА с затратой 3ATP выйдет 3 новых РБФ. Цикл замкнулся - с прикрепления к этим 3 РБФ он начинался.
Подведём итоги
Таким образом в световой фазе фотосинтеза:
6H2O + 6NADP+ + 9ADP + 9Pi =light energy => 3O2 + 6NADP . H2 + 9ATP
В цикле Кальвина до получения ФГА:
3РБФ + 3CO2 + 6NADP . H2 + 6ATP => 6ФГА + 6NADP+ + 6ADP + 6 Pi
В цикле Кальвина после получения ФГА:
5ФГА + 3ATP => 3РБФ + 3ADP + 2Pi (5 C3 => 3 C5 соединений. углерод сохранён)
Вот, смотрите: организм акцептирует 3 атома углерода и выводит из цикла 1 трёх углеродный сахар (ФГА). Вспомним, что на получение всех остальных компонентов в световой фазе было затрачено всего несколько молекул воды! Это дешёвый ресурс для большинства организмов, особенно водных. То есть единственное, что действительно теряется при получении ФГА - это фосфор в его составе. Так что растениям (и не только) нужно черпать неорганический фосфор из внешней среды.
Отступления
№1
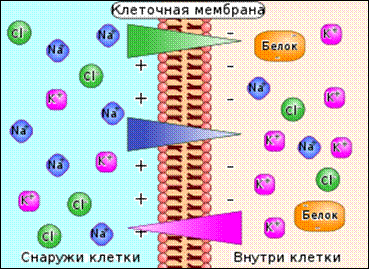 Отступление про эл. хим. градиент:
Отступление про эл. хим. градиент:
Электрохимический градиент - градиент, определяющий направление движения ионов (например, в клетках и компартментах клеток). Он создаётся за счёт различных концентраций (одинаковые ионы идут туда, где их меньше, чтобы создать равновесие) и разности потенциалов (ионы стараются создать заряд равный 0 мВ на мембране). Многие организмы приспособились к этому эффекту и за счёт различных мембранных белков создают нужный себе заряд на мембране (потенциал покоя в нервной системе) или концентрацию определённых ионов (солёная/пресная вода для рыб).
№2
Отступление о различных C - путях: Темновая фаза у разных растений может идти по-разному. Различают как минимум 4 различных пути (названия соответствуют количеству углеродов в ключевых соединениях пути):
C3 путь (так же Цикл Кальвина или Восстановительный пентозофосфатный цикл) - свойствененвсемфотосинтезирующимрастениям. Его и будем рассматривать. Классический, изначальный вариант. Все остальные пути являются надстройками и приспособлениями к конкретным условиям.
C2 путь (фотодыхание) - не путайте с клеточным дыханием. Приспособление фототрофов к особенностям RuBisCO (см. далее).
C4 путь - есть кукурузы, проса. Очень ярко выражен у сахарного тростника. Растения, имеющие такой вариант темнвой фазы, имеют такие анатомические приспособления как редукция мезофилла листа и развитие кранц-обкладки (визуальное увеличение клеток обкладки жилок в листе). Основная цель C4 пути — увеличение концентрации CO2в клетках, в которых протекает C3. Это опять же к проблемам, связанным с RuBisCO (см. далее).
CAM-метаболизм - особый вариант пути фиксации CO2 у растений засушливых местообитаний, гидрофитов и гидатофитами так же в водоёмах с высокой температурой. Он позволяет разделить функционал на дневную и ночную части, так чтобы под палящим солнцем не нужно было открывать устьица в целях дыхания (У водных с этим проще — они могут открывать и днём. Их проблема — содержание в воде доступного CO2и веществ, из которых его можно получить (HCO3-), возрастающая ночью и уменьшающаяся днём).
№3
Отступление о RuBisCO: Рубиско (рибулозо-1, 5-бисфосфат карбоксилаза оксигеназа) - самый распространённый на земле фермент. Он появился тогда, когда появились первые фотосинтезирующие организмы (бактерии). Тогда в атмосфере не было кислорода и рубиско работал отлично. Но потом, когда кислорода стало больше, когда установилась атмосфера земли, оказалось, что у этого фермента есть очень большой минус:
Он не только карбоксилаза, но и оксигеназа. То есть способен к акцепции и CO2, и O2. При акцепции кислорода фотосинтезирующий организм ничего не получает и даже довольно много теряет. Для ослабления этого эффекта создан C2 путь (см. выше). Автотрофы сохранили рубиско, но стали так же добавлять различные надстройки для увеличения концентрации CO2 в строме хлоропласта (это C4 и CAM метаболизмы). В норме углекислый газ просто диффундирует в клетках. Шанс захвата кислорода велик.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|