- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Полисахариды.
Химический синтез полисахаридов является одной из малоразработанных областей химии углеводов. Причина, очевидно, заключается не только в сложности решения задачи, но и в отсутствии заметных потребностей в синтетических полисахаридах: громадное разнообразие природных полисахаридов и их доступность делают существенно более актуальным изучение их строения и свойств.
Целлюлоза построена из остатков моносахаридного типа - глюкопиранозы. Все гликозидные связи имеют β -конфигурацию. Амилоза устроена аналогично, на связи α.
В гиалуроновой кислоте (один из наиболее распространенных полисахаридов соединительной ткани) в цепи чередуются остатки двух разных моносахаридов – D-глюкуроновой кислоты и N-ацетил-D-глюкозамина – со связями β 1 → 3 и β 1 → 4 соответственно. В агарозе, главном гелеобразующем компоненте агара, также чередуются остатки двух моносахаридов: β -D-галактопиранозы и 3, 6-ангидро- α -L-галактопиранозы.
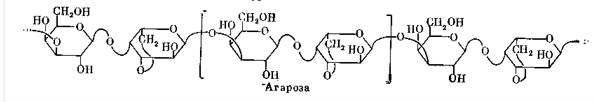
Все перечисленные полисахариды имеют неразветвленные С-цепи и называются линейными.
Замечательная особенность их структур - высокая регулярность. Это значит, что один сравнительно небольшой фрагмент структуры много раз повторяется на всём протяжении цепи, причём других структурных элементов в полисахаридах не содержится. Такой фрагмент называют повторяющимся звеном. Так, например, повторяющееся звено целлюлозы – 1 → 4-связанный остаток β -D-глюкопиранозы, амилозы – 1→ 4-связанный остаток α -D-глюкопиранозы, гиалуроновой кислоты и агарозы – соответствующие дисахаридные остатки.
Понятие повторяющегося звена, даже для таких простейших структур, как целлюлоза, амилоза, гиалуроновая кислота или агароза, не так примитивно, как может показаться на первый взгляд.
Подобная запись не только описывает структуру, но и является прямым выводом из результатов химического анализа этих структур. Так, можно расщепить гиалуроновую кислоту, что практически единственным продуктом будет дисахарид, из чего будет следовать, что именно он и есть мономер.
Более глубокий анализ структур этих полисахаридов может, однако, привести к другим выводам.
Для целлюлозы и амилозы известна конформация цепей.
Для целлюлозы это прямой стержень, в котором каждое следующее звено повернуто на 180º по отношению к предыдущему.
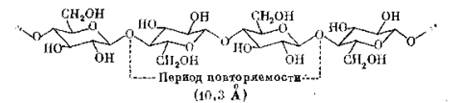
Такая структура действительно строго регулярна и периодична, т. е. сдвиг определенного участка цепи вдоль ее оси приводит к точному наложению на следующий участок – подобно тому, как это имеет место в кристалле.
В этом смысле молекула целлюлозы – одномерный кристалл. Но тогда легко видеть, что такой минимальный участок (шаг цепи) – это не моносахаридный, а дисахаридный остаток. Поэтому с точки зрения конформации цепи повторяющимся звеном в целлюлозе является не остаток глюкозы, а остаток дисахарида целлобиозы.
С амилозой дело обстоит сложнее. Ее молекула – спираль, на одном витке которой помещается точно шесть остатков глюкозы. Как любая правильная спираль, эта система повторяет сама себя, если ее сдвинуть вдоль оси на длину одного витка. В этом смысле повторяющееся звено амилозы представляет собой отрезок цепи из шести моносахаридных остатков, т. е. остаток гексасахарида мальтогексаозы.
Применительно к гиалуроновой кислоте и агарозе дисахаридные фрагменты также не отражают строения конформационных повторяющихся звеньев этих полисахаридов. Это еще один аспект понятия повторяющегося звена. Дисахариды, как уже говорилось, являются продуктами частичного расщепления цепей химическими методами. Поэтому логично. приписать этим полисахаридам структуру из повторяющихся звеньев именно этих дисахаридов.
Т. к. к такой структуре приводит логика химического исследования, соответствующий фрагмент принято называть «химическим» повторяющимся звеном. Точно такую же цепь можно, однако, построить иначе, взяв за основу структуру изомерных дисахаридов.
Многие полисахариды синтезируются клеткой по такой схеме: сначала происходит синтез олигосахарида, а затем его поликонденсация, сшивание в длинные цепи. Такой олигосахарид в биологическом смысле, т. е. с точки зрения путей биосинтеза этого полисахарида, и является истинным мономером полисахаридной цепи. Поэтому такой фрагмент называют «биологическим» повторяющимся звеном. И оно совсем необязательно совпадает с химическим повторяющимся звеном.
Можно, наконец, рассмотреть и еще один – также биологический – аспект понятия о повторяющемся звене, связанный с взаимодействием готовой полисахаридной цепи с другими макромолекулами в живых системах.
Речь в данном случае идет о том, каков минимальный фрагмент цепи, воспринимаемый другими молекулами или системами (назовем их рецепторами) как характерный признак данного полисахарида. Сюда относится широкий круг проблем, таких, как иммунные реакции организма, сортировка макромолекул в клетке и в организме, преодоление клеточных барьеров, метаболизм полисахаридов и т. д.
Рассмотрим лишь один наиболее простой пример: ферментативный гидролиз полисахаридов. Распространенный фермент животных организмов (лизоцим) специфически расщепляет гликозидные связи β 1- 4-связанных остатков N-ацетил-D-глюкозамина в полисахаридных цепях. В частности, он легко катализирует гидролиз полисахаридных цепей хитина.
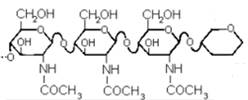
Для того, чтобы фермент мог нормально работать, его рецептору нужно одновременно «чувствовать» участок цепи из шести остатков глюкозамина: тогда он способен выполнять свою функцию – расщеплять четвертую гликозидную связь в этой последовательности. Тогда. «с точки зрения лизоцима», повторяющееся звено в хитине – гексасахаридный фрагмент.
Полисахариды, о которых мы говорили, относятся к числу простейших полисахаридных структур.
Даже неразветвленные полисахариды, построенные из остатков моносахарида одного типа, могут иметь гораздо более сложное строение. Так, например, глюкан овса содержит сопоставимые количества остатков β -D-глюкопиранозы, связанных 1 → 3- и 1 → 4-связями. При этом, эти связи не чередуются правильным образом и не образуют сколько-нибудь значительных блоков из однотипных связей.
Т. е. строение хаотично. Мы, однако, пока не знаем, является ли хаотичность истинной или кажущейся. В самом деле, здесь может быть (хотя отнюдь не обязательно должна быть) упорядоченность высшего порядка, которую мы пока не в состоянии уловить при помощи существующих (относительно грубых) методов исследования.
Пример: статистический анализ страницы текста
Таким образом, мы видим уже два принципа построения полисахаридных цепей:
1правильное чередование (регулярность);
2хаотическое;
3 блочный принцип.
Так устроена альгиновая кислота – полисахарид гелеобразователь избурых водорослей
β -D-маннуроновой кислоты и α -L-гулуроновая кислоты, свяязаны 1 à 4-связями.
А-А-А-А-А-А-А-А-А-А-Б-А-Б-А
Б-А-Б-Б-А-Б-Б-Б-Б-Б
Наконец, сравнительно недавно было установлено, что некоторые полисахаридные цепи могут быть одновременно и нерегулярными, и регулярными, не нарушая при этом незыблемый принцип логики – закон исключительного третьего.
Возьмем один из полисахаридов красных водорослей – порфиран. Он построен из чередующихся остатков D- и L-галактопиранозы (и в этом смысле подобен агарозе). Однако часть остатков D-галактозы превращена в метиловый эфир (по положению 6), а остатки L-галактозы входят в полисахарид частично в виде эфиров серной кислоты по положению 6, а частично в виде 3, 6-ангидропроизводного, как в агарозе. Вариации распределены вдоль цепи хаотически. если порфиран обработать щелочью все остатки L-галактозы становятся одинаковыми: нерегулярность по этим остаткам исчезает.
Дальше можно метилировать (важная реакция для установления структуры). При этом унифицируется все остатки D-галактозы.
Метилирование заведомо регулярного полисахарида (агарозы) приводит к точно такому же (идентичному) метиловому эфиру (из порфиана)
Такая «регулярность-нерегулярность» была обнаружена английским ученым Рисом и названа им «замаскированной регулярностью» или «замаскированной повторяющейся структурой».
Можно полагать, что многие типы полисахаридов имеют достаточно регулярный скелет цепи, в которой некоторые звенья варьируют по типу или (и) конфигурации отдельных заместителей, что сравнительно мало отражается на геометрии макромолекулы в целом. А иногда, наоборот, такие вариации вносят в регулярную структуру определенные, биологически осмысленные нарушения, вызывающие, например, изломы в правильных спиралях. До сих пор мы рассматривали только линейные полисахариды и видели, что даже для простейших биополимеров этого класса, построенных из остатков одного-двух моносахаридов, возможны весьма значительные вариации типов структур, не говоря уже о бесчисленных вариациях конкретных структур внутри каждого типа. Системы, однако, резко усложняются, если учесть существование разветвлений.
Геометрия разветвленных полисахаридов может быть схематически классифицирована следующим образом. Простейшие разветвленные системы содержат одну длинную линейную цепь, к которой присоединены разветвления в виде одиночных моносахаридных остатков или в крайнем случае в виде коротких олигосахаридов.
Так устроен, например, ксилан, выделенный из березы. К регулярной цепи из β 1-4-связанных остатков D-ксилопиранозы присоединены единичные остатки 4-О-метил-D-глюкуроновой кислоты, в среднем один на каждые десять ксилозных звеньев. Такие системы иногда называют «гребнеобразными полисахаридами». 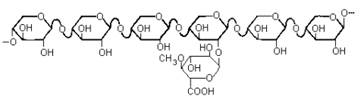
Следующий шаг на пути усложнения структур – полимерный характер боковых цепей, т. е. случай, когда боковые цепи сами являются остатками более или менее высокомолекулярных полисахаридов. Простейшим примером может служить один из полисахаридов бурых водорослей – так называемый растворимый ламинарин.
Его главная цепь построена из β -1-3-связанных остатков D-глюкопиранозы, некоторые из которых несут разветвления в положении 6, а сами разветвления представляют собой тоже регулярные полисахаридные цепи, структурно вполне аналогичные главной.
Изобразим схематически

Боковые полисахаридные цепи, в свою очередь, могут быть разветвлены, а полисахариды, присоединенные к этим боковым цепям, также могут нести разветвления и т. д. Так строятся древовидные структуры высокоразветвленных полисахаридов.
Пример – амилопектин и гликоген
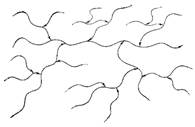
Все цепи построены однотипно (из α -1 → 4). Точки ветвления – построены так же единообразно: боковые цепи присоединены α -1→ 6 гликозидной связью.
И, наконец, последний структурный тип полисахаридов можно было бы назвать ультраразветвленным.
Так устроен галактан одного из видов улиток. К остатку галактозы присоединено два галактозных остатка в положениях 3 и 6. Каждый из этих остатков, в свою очередь, несет по два других галактозных остатка, также присоединенных в положения 3 и 6 и т. д. Таким образом весь полисахарид (а он весьма высокомолекулярен) состоит из сплошных разветвлений.
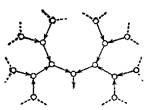
Острый конец – гликозидный гидроксил.
Каждый моносахаридный остаток, за исключением концевых, является узлом ветвления, а понятие главной цепи теряет смысл.
Следует сказать, что между четырьмя названными крайними типами:
1) может существовать бесчисленное множество промежуточных структур;
2)структуры узлов ветвления и полисахаридных цепей внутри одной полисахаридной молекулы вовсе не обязательно одинаковы;
3)полисахаридные молекулы могут быть построены не из одного, а из двух, трех, четырех, пяти, шести, семи и даже восьми типов моносахаридов.
(Подчеркнем, что мы разбираем не просто теоретические возможности, а структурные особенности, встречающиеся в реальных полисахаридах).
Все это создает гигантские, истинно неисчерпаемые возможности вариаций полисахаридных структур и их пространственной организации, далеко превосходящие возможное разнообразие структур каких-либо других типов макромолекул живых систем
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|