- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
ЗОВНІШНІ УМОВИ ДИХАННЯ
Лекція №
Тема: ЗАСВОЄННЯ ВУГЛЕЦЮ І ЕНЕРГІЇ СВІТЛА
ВИНИКНЕННЯ ФОТОСИНТЕЗУ ТА ЙОГО ЗНАЧЕННЯ.
ДИХАННЯ РОСЛИН
ЗНАЧЕННЯ ДИХАННЯ В ЖИТТІ РОСЛИН
План:
1. Фотосинтез
2. Хлоропласти
3. Характеристика світла
4. Особливості фотохімічної реакції
5.Зовнішні умови і фотосинтез
6. Фотосинтез і врожай
7. Дихання рослин
ЗНАЧЕННЯ ДИХАННЯ В ЖИТТІ РОСЛИН
Фотосинтез — це утворення складних органічних речовин у зеленому листку з максимально окислених сполук води і вуглекислоти за допомогою світла і хлорофілу.
Фотосинтез — єдине джерело вільного кисню на Землі, трансформатор світлової енергії Сонця. Незвичайна складність цього процесу зумовлена поступовим розвитком і вдосконаленням фотосинтезу в ході еволюції організмів.
Зелені рослини належать до автотрофних організмів, що утворюють в процесі фотосинтезу необхідні для життя органічні речовини з мінеральних сполук вуглецю, азоту, сірки, фосфору та інших елементів. Під час фотосинтезу рослини засвоюють вуглець із зовнішнього середовища для утворення органічних речовин, що становлять 95 % сухої маси врожаю, і запасають у них енергію, яку потім використовують як рушійну силу всіх життєвих процесів не тільки в зелених рослин, айв усіх представників живого світу. Рослина — цілісний організм, в якому функції живлення (листкове і кореневе) ніяк не замінюються і не виключаються. Здебільшого рівень мінерального живлення і водопостачання бувають мінімальні; зміни їх завдяки обробітку ґрунту, поливанню, внесенню добрив є досить ефективним і доступним засобом впливу на формування врожаю через підтримання фотосинтезу.
Людина з’явилась на Землі серед більш пізніх представників гетеротрофних організмів, тому її життя цілком залежить від фотосинтезу рослин: вона дістає їжу тільки від рослин або безпосередньо у вигляді плодів, бульб, коренеплодів, або через тварин у вигляді м’яса, молока, яєць.
Провідне значення фотосинтезу може характеризуватись такими даними. У період інтенсивного росту добові прирости сухої маси становлять в середньому 150 кг, а в кращому варіанті — 500 кг на 1 га. При цьому корені засвоюють у вигляді іонів 2 кг азоту, 0,5 кг фосфору, 4 кг калію і 4 кг інших елементів. За цей самий час листя засвоює 1000 кг С02. При середній урожайності коренів цукрових буряків 300 ц з 1 га рослини у вегетаційний період засвоюють 150 кг азоту, 30 кг фосфору, 160 кг калію і 4200 кг вуглецю, що відповідає поглинанню 20 т СО2. В урожаї акумулюється 167,2 ГДж енергії. Це треба мати на увазі, обґрунтовуючи мету і завдання землеробства, яке є системою використання основної функції зелених рослин — фотосинтезу.
Усі заходи цієї системи спрямовані на підвищення продуктивності фотосинтетичного апарату рослини-листка.
2. ХЛОРОПЛАСТИ
Фотосинтезуючими елементами клітин листка є хлоропласти — пластиди, що містять зелений пігмент хлорофіл. Хлоропласти, беручи участь у фотосинтезі, постачають хімічну енергію для живих організмів. Без хлоропластів не змогли б жити ні рослини, ні тварини, які безпосередньо або опосередковано живляться рослинами.
Форма, розмір і розподіл хлоропластів сталі для кожного з видів рослин. У клітинах вищих рослин хлоропласти можуть бути сферичної, яйцевидної або дископодібної форми. У водоростей роль хлоропластів виконує хроматофор, що має вигляд сітки, спіральної стрічки або зірчастої пластини і завжди один.
Розмір хлоропластів у вищих рослин 4-6 мкм, а хроматофор у водоростей дорівнює розміру клітини. В основному розмір хлоропластів зумовлений світловим режимом життєвого середовища рослин. У світловитривалих рослин вони значно дрібніші, ніж у тіневитривалих. Розподіляються хлоропласти по цитоплазмі рівномірно, але буває й скупчуються біля ядра і клітинних оболонок — залежно від розміщення поверхні листка відносно напряму сонячного променя.
Кількість хлоропластів, так само як і їхня форма, для кожного з видів рослин стала. У вищих рослин кількість хлоропластів збільшується завдяки поділу кожного хлоропласта навпіл перетяжкою посередині. Процес поділу одного хлоропласта відбувається за 8 днів. У водоростей є тільки один хлоропласт.
Хлоропласти містять у складі сухої маси білків 35-55%, в основному у вигляді ліпопротеїдів і ліпідів 20-30%, у вигляді жирів, восків, фосфатидів, стеринів та їхніх компонентів: холіну, інозиту, гліцерину, етаноламіну, серину. Вуглеводи містяться у вигляді крохмалю та фосфатів сахарів: тріоз, тетроз, пентоз, гексоз і гептоз. Кількість їх дуже нестала. Крім того, в хлоропластах є філохінони і токофероли (вітаміни К і Е), цитохроми, метали (залізо, мідь, марганець, цинк), окислювально-відновні ферменти гідролази та ін.
Структура хлоропластів гетерогенна. Зовні хлоропласт має подвійну мембрану з малесенькими отворами. Під нею розміщена строма (матрикс), пронизана ламелами — пластинками, утвореними складною системою внутрішніх мембран. У певних місцях хлоропласта ламели переходять у дископодібні замкнені мішечки — тилакоїди, зібрані в щільні стовпчики — грани. Грани — плоскуваті тільця, що мають форму пластинок або дисків, покладених одне на одного у вигляді щільних стовпчиків, перпендикулярних до поверхні хлоропласта. Розмір гран — від 0,3 до 1,7 мкм. Грани з’єднуються одна з одною за допомогою мембран або трубочок. У хлоропласті може бути 40-60 гран середнього розміру. Грани містяться в менш щільному білковому матриксі. У водоростей гран немає.
У гранах хлорофіл буває у вигляді мономолекулярного шару (рис. 11).
Хлорофіл — пігмент зеленого кольору, властивий клітинам тільки зелених рослин. Без нього листок не може здійснити синтез речовин на світлі в процесі фотосинтезу.
У рослинах виявлено різні хлорофіли, їх позначають малими літерами латинського алфавіту: а, в, с, d. Хлорофіл в відрізняється від хлорофілу а тим, що в його структурі одна метильна група замінена альдегідною і він має жовтий відтінок, тимчасом як хлорофіл а — синій. Решта — це суміш хлорофілів.
Великих успіхів у галузі виділення, очищення, встановлення структури і вивчення біохімічних реакцій пігментів листка біохімікам вдалося досягти завдяки геніальному своєю простотою методові хроматографічного адсорбційного аналізу, розробленому в 1903 р. російським ученим М. С. Цвєтом.
Принцип цього методу покладено в основу нових методів хроматографії: розподільчої хроматографії на папері, тонкошарової і газової.
Властивості хлорофілу нині вивчено досить докладно. В основу вивчення цих властивостей покладено геніальні праці М. В. Ненцького, К.А. Тімірязєва, М. С. Цвєта, Р. Вільштеттера і Г. Фішера.
Хлорофіл — це складний ефір двоосновної хлорофілінової кислоти і двох спиртів: метилового і високомолекулярного ненасиченого спирту фітолу, який надає хлорофілові ліпідних властивостей, що проявляються в його розчинності в жирових розчинниках і наявності воскоподібної консистенції. Гідрофільні властивості зумовлені ковалентними зв’язками між магнієм і азотом, а також наявністю циклопентанового кільця. Отже, молекула хлорофілу з довжиною поперечного перерізу 22,5 нм має водночас гідрофільні і гідрофобні властивості, що дає їй змогу міцно сполучатися з білком і ліпідом. Слід підкреслити, що В.М. Любименко (1921) був першим ученим, який висловив передбачення про зв’язок хлорофілу з білком і назвав його зеленим білком. За даними хімічного аналізу, натуральний хлорофіл містить 63 % білка, 21 % ліпоїдів, 8 % пігментів і 2 % інших речовин. Додаткові зв’язки між магнієм і азотом зумовлюють зв’язок з білком, а ковалентні — зелене забарвлення і поглинання червоних променів. Подвійні зв’язки забезпечують поглинання хлорофілом синьо-фіолетових променів і підвищують його хімічну активність. Отже, хлорофіл — єдиний пігмент, спільний для всіх фотосинтезуючих організмів. Тому тільки хлорофіл а здатний бути донором енергії безпосередньо для фотосинтетичної реакції, а всі інші пігменти передають йому поглинуту енергію.
Крім хлорофілів, у клітинах листка у невеликій кількості (0,07-0,2% сухої маси) є каротиноїди. Це сполуки, розчинні в жирах і органічних розчинниках, з довгим ланцюгом спряжених подвійних зв’язків. Поглинаючи короткохвильові промені, каротиноїди виконують роль екрана, що захищає хлорофіл і клітину від їхньої згубної дії.
3. ХАРАКТЕРИСТИКА СВІТЛА
Промениста енергія, така необхідна для рослин,— це енергія електромагнітних коливань, що виникають в результаті коливальних рухів електронів, збуджених атомів і молекул. Згідно з сучасними уявленнями, промениста енергія випромінюється і поширюється не безперервно, а окремими неподільними порціями — квантами, або фотонами, величина яких визначається довжиною хвилі або частотою:

де А — стала Планка, що дорівнює 6,55-10~37 ерг-с; V — частота; с — швидкість, що дорівнює 300000 км/с; А. — довжина хвилі.
Згідно з законом Ейнштейна, інтенсивність фотохімічних процесів залежить не від абсолютного поглинання енергії безвідносно до її квантової природи, а від кількості поглинутих квантів певної величини. При одному й тому ж поглинанні енергії кількість поглинутих квантів зростає разом із довжиною хвилі.
Спектр сонячного світла щодо фізіологічної характеристики обмежується довжинами хвиль від 300 нм і менше до 6000 нм і більше. Фотосинтетично активна радіація (ФАР), яка поглинається пігментами листка і забезпечує нормальний ріст і розвиток рослин, становить 300—750 нм.
Короткі ультрафіолетові промені з довжиною хвилі <300 нм згубні для всіх живих організмів Землі. Це вітацидна радіація. Ультрафіолетові промені з довжиною хвилі 300—400 нм, які поглинаються на 99 %, є росторегулюючими променями, що запобігають надмірному витягуванню рослин. Під дією цієї радіації у рослин змінюється хімічний склад, підвищується вміст білків, вітамінів.
Синьо-фіолетові промені з довжиною хвилі 400-500 нм, які поглинаються хлорофілом, каротиноїдами, ферментами та іншими компонентами клітини, здебільшого затримують перехід рослини до цвітіння, сприяють синтезу білків і впливають на хімічний склад рослин.
Зелені промені з довжиною хвилі 500-600 нм, які поглинаються на 75 %, мінімально активні, фізіолого-біохімічні процеси дуже загальмовані, і вирощування рослин нераціональне через велику витрату променистої енергії.
Оранжево-червоні промені з довжиною хвилі 600-700 нм — максимально активні і поглинаються тільки хлорофілом на 95 %. У цих променях з максимальною швидкістю здійснюється фотосинтез, утворення листків, формування коренеплодів, цибулин, стеблових потовщень, перехід рослин до цвітіння, фотоперіодичний процес тощо. При цьому буває найвищий урожай.
Інфрачервоні промені з довжиною хвилі 700-1000 нм не поглинаються або дуже мало поглинаються (15 %) пігментами і тканинами рослин. Фізіологічного значення вони практично не мають, і тому їхнє випромінювання називають абіотичною радіацією.
Далекі інфрачервоні промені з довжиною хвилі 1000-6000 нм, які поглинаються на 90 % в основному водою, пігментами і цитоплазмою, визначають тепловий режим рослинних тканин. Вони впливають на швидкість і напрям фізіолого-біохімічних процесів відповідно до температури навколишнього середовища. При низьких температурах повітря (<20°С) вони зігрівають тканини, а при високих температурах (>30 °С) негативно впливають на обмін речовин.
Видиму сонячну радіацію з довжиною хвилі 400-720-нм рослини поглинають неоднаково. Поглинання видимої радіації рослиною зростає із зменшенням широти місцевості, із збільшенням прозорості атмосфери, з висотою над рівнем моря, з онтогенезом (індивідуальним розвитком рослин) і переважно у деревних.
При вирощуванні рослин на червоному світлі або в темряві вони набувають вигляду етіольованих: у них забарвлення біле або світло-жовте, стебла дуже витягнені, листя недорозвинене, покривні механічні і провідні тканини розвинені мало. Явище етіоляції використовується для вирощування деяких овочів: спаржі, салату-ромену, капусти та ін. Етіольовані рослини на сонячному світлі швидко зеленіють завдяки утворенню хлорофілу.
Для фотосинтезу, крім води і світлової енергії, потрібен вуглекислий газ, якого в атмосфері в середньому 0,03 %. Максимальна кількість його в ґрунтовому повітрі — до 5%. Завдяки градієнтові активності молекул вуглекислий газ накопичується на поверхні ґрунту, звідки конвекційними повітряними потоками піднімається вгору, в атмосферу. У зв’язку з тим, що вуглекислий газ має більшу густину, ніж повітря, він опускається в низькі місця на поверхні Землі. А тому мінімальну кількість його (0,006 %) зафіксовано високо в горах.
Вуглекислий газ рослини поглинають листками величезною поверхнею клітин паренхіми, що вимірюється гектарами. Пропускаючи крізь листя повітря, рослина просто-таки виловлює молекули вуглекислого газу.
Щоб синтезувати 4 кг глюкози, рослина поглинає 3 м3 вуглекислого газу із 10 000 м3 повітря, з яким треба зіткнутися кожній фотосинтезуючій клітині листків. Визначено, що кожні 200 років весь атмосферний вуглекислий газ пропускається крізь рослини і весь атмосферний кисень оновлюється ними кожних 2000 років. Без рослин в атмосфері не залишилось би кисню і життя на Землі було б практично неможливе.
4. ОСОБЛИВОСТІ ФОТОХІМІЧНОЇ РЕАКЦІЇ
Фотосинтез — процес перетворення світлової енергії в хімічну за допомогою хлорофілу і використання цієї енергії для синтезу органічних речовин, що утворюються з вуглекислого газу і води. Схематично цей процес можна зобразити таким сумарним рівнянням:
СО2 + Н2О= СпН2пОп + 6О2
Естонський учений Ф.І. Гротгус у 1817 р. охарактеризував хімічно активним тільки те світло, яке поглинається реагуючим середовищем. Через 58 років К. А. Тімірязєв установив прямий зв’язок між кількістю продукту, добутого при певній фотохімічній реакції, і кількістю поглинутої світлової енергії. Фотохімічна дія світла полягає у збудженні атомів або молекул реагуючої речовини внаслідок поглинання світлових квантів, або фотонів. Співвідношення між поглинутою енергією і кількістю прореагованої речовини виражається законом фотохімічної еквівалентності: кожна молекула, що реагує під дією світла, поглинає один світловий квант, або фотон. Енергія кванта перебуває в оберненій залежності від довжини хвилі світла.
Фотосинтетична одиниця — це система з кількох сотень молекул хлорофілу а, що збирають енергію світлових квантів і передають її фотохімічному активному центру, де відбувається первинне розділення окислювально-відновного потенціалу і перенесення електронів на дві фотохімічні системи. Ці системи, взаємодіючи одна з одною, беруть участь в асиміляції вуглекислого газу і виділенні кисню з використанням восьми квантів світла.
Виходячи з усього сказаного, слід зазначити, що процес фотосинтезу складається з двох видів реакцій: світлових і темнових. Світлові реакції, або фотохімічні, відбуваються тільки на світлі, оскільки в них енергія світла перетворюється в енергію фосфатних зв’язків. Темпові реакції, або біохімічні, відбуваються незалежно від освітлення, тому що в біосинтезі органічних речовин бере участь хімічна енергія, яка утворилася внаслідок перетворення сонячної енергії у світлових реакціях.
5. ЗОВНІШНІ УМОВИ І ФОТОСИНТЕЗ
Фотосинтез не автономний процес, він регулюється рослиною і сам впливає на інші фізіологічні функції рослини. Тому для розуміння фотосинтезу як фізіологічного процесу необхідно визначити залежність його від різних зовнішніх і внутрішніх факторів і вивчити взаємозв’язок фотосинтезу та інших процесів життєдіяльності рослин.
Світло — головна умова для здійснення фотосинтезу, який починається вже при мінімальній інтенсивності світлової енергії (гасова лампа, вечірня зоря). Із збільшенням інтенсивності світла до 1/3 повного сонячного освітлення швидкість фотосинтезу зростає, потім починає відставати, а при освітленні 250 000 ерг/см2-с настає світлове насичення фотосинтезу. У більшості сільськогосподарських культур світлове насичення буває в межах 85-220 000 ерг/см2-с. При освітленні рослини променями полудневого сонця хлорофіл завдяки зеленому забарвленню пропускає більш інтенсивні в цей час жовто-зелені промені і тим самим захищає хлоропласт від перегрівання. При розсіяному світлі хлорофіл поглинає тим більше його (%), чим слабше освітлення. Така залежність свідчить про пристосувальний характер зеленого забарвлення хлорофілу, який виконує роль поглинача сонячної енергії лише для фотосинтезу, і не приймає надлишку її.
У природних умовах фотосинтез рослин здійснюється звичайно при змінному освітленні. Рослини на відкритих місцях витримують високу напруженість світла і називаються світловитривалими. У них фотосинтез відбувається при високій температурі прямої сонячної радіації і посиленому водообміні. Рослинам густих насаджень, що ростуть під покровом лісу, в печерах, у глибинах вод, дістається світло дуже малої інтенсивності. Ці рослини називають тіневитривалими. У них фотосинтез відбувається при зниженій температурі, розсіяному світлі і слабкому водообміні.
До високої напруженості світла рослини пристосовуються завдяки значному зменшенню відносної кількості хлорофілу в і ксантофілів. Світловитривалість виражається помітним зменшенням вмісту зелених пігментів і відповідним збільшенням каротиноїдів. У світловитривалих рослин часто спостерігається посилений розвиток асимілюючої тканини палісадної паренхіми. Паренхіма в них складається з кількох шарів клітин меншої величини, ніж у тіневитривалих рослин; кількість продихів на одиниці поверхні значно більша, а тому СО2 тут швидше проникає всередину листка. У тіневитривалих рослин епідерміс утворює сочевицеподібні вирости, які концентрують світло. Ці рослини містять більше хлорофілу і їхні хлоропласти більші за розміром. Тіневитривалі рослини на відкритому місці пригнічені і краще розвиваються при зменшенні загальної кількості світла, через те що хлорофіл на розсіяному світлі поглинає тим більше енергії, чим менша його інтенсивність.
Встановлено, що на інтенсивному світлі у рослин утворюється більше вуглеводів, а при малій інтенсивності його синтезується більше азотистих сполук. У посівах рослин через самозатінення інтенсивність світла незначна. Це часто негативно впливає на врожай. Тому рослини, відповідно до умов, в яких вони ростуть, пристосовуються до певної інтенсивності світла, і порушення світлового режиму негативно позначається на житті їхнього організму.
Залежність швидкості фотосинтезу від кількості вуглекислого газу, температури і води, так само як і залежність від дії світла, виражається логарифмічною кривою. Для вуглекислого газу крива обмежена концентраціями 0,006-1,5%, для температури амплітуда ширша — від мінус 15°С до +85°С (для термофілів 90°С) з оптимумом 25-30°С. Водний дефіцит коливається в межах 5—60 %.
Важливі елементи мінерального живлення — азот, фосфор, сірка і магній — є будівельним матеріалом для фотосинтетичного апарату; інші елементи — залізо, калій, хлор, мідь, натрій тощо, які не входять до складу хлоропластів, впливають на накопичення хлорофілу, а отже, й на фотосинтез. Азот, як складова білка і хлорофілу, посилює їхній синтез, забезпечує синтез органічних кислот і амінокислот. Калій, рубідій і цезій, змінюючи колоїдний стан цитоплазми, регулюють інтенсивність фотосинтезу, накопичення хлорофілу, вуглеводів та білків. Фосфор входить до складу фосфорильованих сполук, що беруть участь в асиміляції вуглекислого газу, регенерації пентозофосфатів і в акумулюванні хімічної енергії в фосфатних зв’язках АТФ. При нестачі магнію, заліза, цинку та міді дуже зменшується асиміляція вуглекислого газу і проявляється хвороба рослин — хлороз. Надмір натрію і хлору гальмує фотосинтез і знижує врожай.
6. ФОТОСИНТЕЗ І ВРОЖАЙ
У нормальних умовах величина господарських урожаїв тісно пов’язана з біологічними врожаями, що являють собою суму добових приростів сухої загальної біомаси на 1 га посіву протягом вегетаційного періоду. Звичайно розміри добових приростів сухої маси врожаю коливаються від нуля (на початку і наприкінці вегетаційного періоду або за несприятливих умов) до 150-300 і навіть 500 кг на 1 га (при найбільшому розвитку листя і в сприятливі для фотосинтезу періоди вегетації). У посівах, що перебувають у різних умовах, площа листків може збільшуватися з різною швидкістю. Через різну тривалість вегетаційних періодів у різних рослин період «роботи» листків також різний. Щодо цього відмінність посівів можна характеризувати поняттям «фотосинтетичний потенціал посіву», що виражається сумою площ листків (у м2) на 1 га за добу протягом вегетаційного періоду. Цей показник коливається у рослин помірної зони в межах 50-500 м2 на 1 м2. У формуванні врожаю важливою є продуктивність фотосинтетичної дії 1 м2 площі листків. Вона може коливатись від 4 до 6 г вуглекислого газу за 1 шд при добрих умовах і спадати до часток грама при поганих. Звичайно інтенсивність фотосинтетичної роботи листків характеризується показником чистої продуктивності фотосинтезу, що складається з кількості загальної суми біомаси, утвореної рослиною протягом доби в розрахунку на 1 м2 листкової поверхні. Цей показник за вегетаційний період коливається від нуля і навіть від’ємних значень до 15—18 г на 1 м2 за добу.
Формування врожаю — процес не тільки кількісний, а й якісний. У ньому весь час змінюється живлення, співвідношення між різними його видами, використання речовин, що утворюються в процесі живлення. Спершу переважає ріст вегетативних органів, а потім запасаючих і репродуктивних. При інтенсивному загальному рості і при великій масі біологічного врожаю можна мати і дуже високі, і низькі, і навіть мізерні господарські врожаї. А тому важливо, щоб у відповідні періоди росту розподіл утворюваних і накопичуваних поживних речовин і асимілятів був найбільш сприятливим для формування не тільки загального біологічного, а й господарського врожаю. Для цього треба всі агротехнічні заходи (внесення добрив, поливання, обробіток) спрямувати на підтримання оптимальних умов, які б забезпечили найкращий ріст фотосинтетичного апарату — площі листків; найбільшу тривалість активної роботи листків протягом кожної доби вегетаційного періоду; найвищу інтенсивність фотосинтезу і сум денного засвоєння вуглекислого газу; високу чисту продуктивність фотосинтезу і високі добові прирости сухої речовини; найбільш високий коефіцієнт господарської ефективності фотосинтезу, що виражається відношенням маси господарської частини врожаю до маси біологічного врожаю.
7. ДИХАННЯ РОСЛИН
ЗНАЧЕННЯ ДИХАННЯ В ЖИТТІ РОСЛИН
Дихання є дуже важливим фізіологічним процесом, під час якого відбувається виділення енергії, необхідної для нормального функціонування рослин, а також утворення ряду органічних сполук, потрібних для синтезу компонентів клітини.
Інтенсивність дихання і характер його окислювальної системи зумовлюються систематичними і екологічними групами. Найвища інтенсивність дихання у бактерій і грибів, а також у клітин твірної меристеми коренів вищих рослин.
Дихання залежить від екологічних умов росту рослини. У світловитривалих рослин воно інтенсивніше, ніж у тіневитривалих. Підвищена інтенсивність дихання властива також арктичним рослинам, особливо при низьких температурах. Крім того, їм властива підвищена метаболічна активність у зв’язку з перебігом всього циклу розвитку за короткий період вегетації. Це також стосується високогірних рослин, які адаптувалися до низького парціального кисневого тиску. Ця особливість притаманна і тваринам високогірних районів. У них дуже збільшена кількість крові в організмі і вміст у ній еритроцитів. Знижена інтенсивність дихання властива сукулентам через масивність тканин, поверхня яких, вкрита товстим шаром кутикули, мало проникна для газів.
Структура тканин та їхня біохімічна активність помітно змінюються з віком, що позначається на ступені дихального процесу. Інтенсивність дихання сухого насіння дуже слабка, але після намочування різко активізується, що пов’язано з утворенням у зоні росту нових меристематичних клітин. Через кілька днів після проростання дихання досягає максимуму, а потім спадає внаслідок старіння цитоплазми новоутворених клітин і зменшення відносного значення меристематичних клітин. У багатьох видів рослин виявлено посилене дихання у старіючих листків (жита, сорго, вівса) внаслідок відмирання клітин, незбалансованості окислення субстратів дихання, що веде до тимчасового посилення поглинання кисню та виділення вуглекислого газу. В період цвітіння рослина дихає інтенсивно, особливо її репродуктивні органи (тичинки, маточка). При дозріванні активність дихання спадає, особливо наприкінці повної спілості, що супроводжується висиханням зерна. Зниження дихання триває і в період післязбирального дозрівання, пов’язаного водночас із підвищенням схожості й енергії проростання зерна.
Ритмічність дихання рослин спостерігається не тільки в онтогенезі, а й протягом доби. Опівдні дихання в листків цукрових буряків максимальне, опівночі — мінімальне. Це результат філогенетичного пристосування до закономірної зміни зовнішніх умов. Зміна дихання в онтогенезі має не тільки кількісний, а й якісний характер. У молодих тканинах (пшениця, рицина, соняшники, помідори) переважає гліколітичний шлях окислення глюкози (анаеробний), а в старіючих — гексозомонофосфатний. Провідна роль в активації молекулярного кисню при проростанні належить ци- тохромоксидазі, а потім із ростом проростка переходить до флавопротеїнових оксидаз. Під час утворення насіння на перше місце знов виходять цитохромоксидази. Отже, зміни в інтенсивності і характері дихання протягом онтогенезу рослини тісно пов’язані з особливостями обміну речовин у рослині залежно від філогенетичного розвитку і екологічної групи.
ДИХАННЯ — ОСНОВА ОБМІНУ РЕЧОВИН І ЕНЕРГІЇ
Дихання — окислювально-відновний процес вивільнення хімічної енергії з органічних речовин — є ніби фокусом, що об’єднує всі процеси життєдіяльності рослини, в тому числі білковий, вуглеводний та ліпідний обміни. При окисленні субстрату дихання утворюється багато проміжних сполук, які можуть бути вихідними речовинами для різних біосинтезів. Наприклад, трифосфогліцериновий альдегід, що утворюється при окисленні глюкози, може бути вихідним продуктом для синтезу гліцерину, складової молекули жиру. Піровиноградна кислота як продукт окислення трифосфогліцеринового альдегіду в окислювальному декарбоксилюванні, сполучаючись з КоА, перетворюється в ацетил-КоА, який є основним будівельним матеріалом при біосинтезі жирних кислот. У реакціях амінування піровиноградна кислота є вихідною речовиною для біосинтезу таких амінокислот, як аланін, фенілаланін, тирозин, серин та ін. Фосфоенолпіровиноградна кислота і еритрозо-4-фосфат, що утворюється при апотомічному окисленні, конденсуючись, замикаються в шестичленне кільце шикімової кислоти, яка є попередником ряду амінокислот, а через них ауксинів. Аналогічно утворюються кільця фенолів, лігніну, антоціанів та ін.
Рослина дихає при безпосередній участі різних вітамінів, які є коферментами окислювально-відновних ферментів. У цьому виявляється тісний взаємозв’язок процесів асиміляції і дисиміляції в живому організмі.
Крім того, в процесі дихання організм дістає енергію для своєї життєдіяльності. Гетеротрофи поглинають готові органічні речовини, які, окислюючись у процесі дихання, забезпечують організм пластичними речовинами і енергією. А тому гетеротрофи здатні здійснювати обмежену кількість біосинтезів. Автотрофи мають більше біосинтезів, оскільки крім дихання вони використовують сонячну енергію і синтезують різноманітні органічні речовини з неорганічних: води, вуглекислого газу, аміаку і нітратів. Енергія, що вивільняється як в результаті окислювального, так і фотосинтетичного фосфорилювання, фіксується у вигляді макроергічних зв’язків — трифосфатів.
Крім біосинтезів, хімічна енергія дихання йде на підтримання структури цитоплазми, процесу росту, на надходження і рух поживних речовин тощо. Тонус клітини, що дає змогу їй зберегти форму і впорядкованість процесів, потребує постійного споживання енергії від АТФ. Надходження речовин у клітину з навколишнього середовища проти градієнта концентрації є активним процесом, що відбувається із споживанням енергії. Якщо дихання порушується, то поглинання і рух речовин крізь клітинну оболонку здійснюється за законами дифузії. Виділення енергії при інтенсивному диханні змінює мікроклімат рослини. Підвищення температури в суцвіттях ароїдних приваблює комах. Ріст пролісків супроводжується підвищенням температури, а тому сніг навколо них тане. І нарешті, така важлива властивість рослини, як її стійкість проти патогенів, також пов’язана з диханням. Ця властивість особливо; проявляється під час зберігання зерна, плодів, коренеплодів, бульб, цибулин та ін. Надійно зберігати їх можна тільки в умовах, що забезпечують нормальний дихальний газообмін.
В умовах зберігання дихання залежить від біологічних особливостей об’єкта, що зберігається, температури і газового складу атмосфери. Чим нижча температура, тим нижчий рівень дихання і менші втрати поживних речовин. Низька температура гальмує розвиток патогенів і знижує транспірацію, що має важливе значення для зберігання соковитих плодів. Зберігання картоплі при температурі нижчій, ніж 4° С, посилює гідроліз крохмалю і погіршує смакові якості. Аналогічне явище спостерігається у зберіганні цитрусових при температурі, нижчій ніж 6°С. Для капусти оптимальною є температура мінус 1°С, а для зберігання зерна вирішальним фактором, крім температури, є вода, яка може активізувати дихання через набухання і виділення при цьому енергії.
Склад атмосфери, як і низька температура, по-різному впливає на зберігання. Так, знижений вміст кисню і підвищений вуглекислого газу позитивно впливає на яблука» груші, моркву, але негативно — на капусту і картоплю.
Отже, дихання рослинного організму — різнобічний процес, що охоплює життєдіяльність рослини не тільки в онтогенезі, а й у філогенезі (розвиток виду).
АНАЕРОБНЕ ОКИСЛЕННЯ
Запасні клітинні речовини (крохмаль, білки, жири та ін.) піддаються різним перетворенням, перш ніж їх буде використано для дихання. Вони розщеплюються на більш прості продукти піврозпаду. Мономери, що утворюються, клітина може використати у вигляді різних похідних. Рослинна клітина має каталітичні системи, за допомогою яких глюкоза може окислюватись: по-перше, прямим окисленням вільного сахару за допомогою фламінової глюкооксидази, по-друге, використанням один раз фосфорильованої глюкози в процесі апотомного або гексозомонофосфатного шляху, по-третє, використання двічі фосфорильованої гексози в гліколізі, або шляху ЕМП (Ембден — Майєргофф — Парнас) .
Окислення вільної глюкози на початкових етапах дихання веде до утворення 2-кетоглюконової кислоти, що розпадається на дві тріози, які на другому етапі окислюються до вуглекислого газу і води. Обидва етапи відбуваються у аеробних умовах.
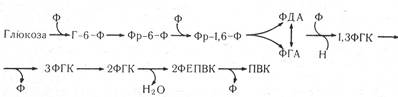
Рис. 17. Гліколіз (шлях ЕМП).
Двічі фосфорильована гексоза використовується в два етапи: 1) анаеробний (анаеробне окислення), коли глюкоза після фотосинтезу або біосинтезу перетворюється у дві тривуглецеві молекули піровиноградної кислоти, і 2) аеробний (аеробне окислення), в результаті якого піровиноградна кислота окислюється з утворенням вуглекислого газу і води. За своєю хімічною природою процеси анаеробного розщеплення глюкози у рослинній клітині близькі до процесів у м’язах тваринних організмів і так само названі гліколізом, або шляхом ЗМП (рис. 17). Своїми працями Л. Пастер, Є. Пфлюгер, В. Пфеффер, В. І. Палладій, С. П. Костичев довели, що початковий етап нормального кисневого дихання — це анаеробний розпад молекули моносахару. У розробці хімізму гліколізу взяли участь видатні вітчизняні і зарубіжні біохіміки і фізіологи Л. А. Іванов, А. Н. Ле- бедєв, А. Гарден, К. Нейберг, Г. Ембден, О. Майєргофф, Я. Парнас та інші вчені, які встановили, що процес гліколізу складається з ряду взаємозв’язаних, суворо послідовних реакцій, каталізованих відповідними ферментами.
АЕРОБНЕ ОКИСЛЕННЯ
Подальшим етапом дихання є аеробне окислення піровиноградної кислоти до вуглекислого газу і води. У послідовних реакціях окислення піровиноградної кислоти спостерігається активізування водню субстрату і перенесення його і електронів на кисень. Цей комплекс перетворень називається лимоннокислим циклом, або циклом трикарбонових кислот, або циклом Кребса. Перші праці про вплив органічних кислот на дихання належать А. Сент-Дьордьї, який виявив, що дихання посилюється від внесення в живильне середовище чотирьох кислот — янтарної, фумарової, яблучної, щавлевооцтової. У 1937 р. Г. Кребс запропонував схему лимоннокислого окислення піровиноградної кислоти через трикарбонові кислоти. Схемуназвано його ім’ям. Виявилось, що цикл Кребса є тим спільним руслом, яким і відбувається остаточне окислення субстратів дихання — вуглеводів, білків та ліпідів — у всіх живих організмів (рис. 18).
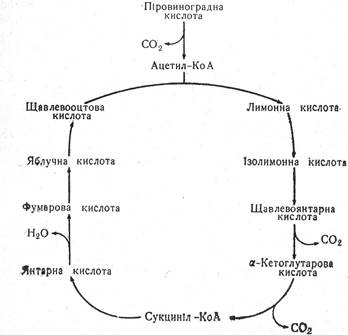
Рис. 18. Цикл Кребса.
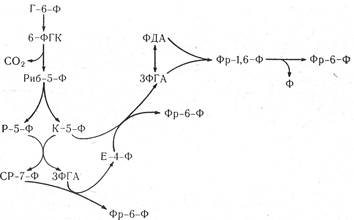
Рис. 19. Гексозомонофосфатний шлях.
Повне окислення глюкози до СО2 відбувається за шість обертів циклу. Апотомічне окислення має ряд переваг порівняно з гліколізом і циклом Кребса: перед тим як вивільнити СО2, глюкоза проходить три проміжні стадії перетворення (6-фосфоглюконалактон, 6-фосфоглюконат і рибульозо-5-фосфат), тимчасом як у гліколізі дегідрування спостерігається тільки у двох реакціях, а в гліколізі і циклі Кребса — в шести.
ЗОВНІШНІ УМОВИ ДИХАННЯ
Для структури дихальної системи і інтенсивності дихального газообміну велике значення мають особливості організму і умови зовнішнього середовища. Вона є не тільки кінцевим продуктом окислення, а й реагентом окремих реакцій дихання. Кількість води, потрібної для дихання, визначається фізіолого-біохімічним рівнем організму. В сухому насінні дихання знижене, через те що немає водяної фази. Збільшення вологості не тільки посилює дихання, а й змінює реагування процесу на температуру. Максимум температури для більшості рослин перебуває в межах 45-55°С, а мінімум — близько 0°С (какао 8°С, бруньки зимуючих багаторічних мінус 25 °С).
На інтенсивність дихання дуже впливає коливання температури, як правило, посилюючи його. Пристосування дихання до коливання температури проявляється в якісних змінах окремих ферментів (на початку розвитку у плодів яблуні переважає флавопротеїноксидазна система, під час наливання — цитохромоксидазна, у спілих плодів — поліфенолоксидазна).
Найефективніше впливає на дихання короткохвильова радіація, особливо ультрафіолетові промені. Отже, вплив світла на дихання визначається не тепловою енергією, а специфічним впливом певних ділянок спектра.
Мінеральні речовини впливають на дихання різними шляхами. Іони металів входять до складу ферментів (залізо — в активну групу каталази, пероксидази, цитохромоксидази; молібден — нітратредуктази, ксантиноксидази; мідь — поліденолоксидази, аскорбіноксидази), можуть бути активаторами ферментів (магній — фосфотази, енолази, фосфоглюкомутази, кінази; калій — фруктокінази, піруваткінази, ацетилази КоА) та їхніми інгібіторами (солі важких металів чинять неспецифічне інгібування ферментів).
Інтенсивність дихання змінюється залежно від газового складу. Клітини надземних органів рослин перебувають у газовому середовищі, де об’єм кисню становить п’яту частину. Підземні органи зазнають різних коливань кисню, які залежать від структури і во
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|