- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Комментарии 2 страница

Основные типы секреции: мерокриновая, апокриновая, голокриновая
Железистые эпителии имеют выраженную секреторную функцию. Клетки, образующие железистый эпителий, называют грандулоцитами. Они синтезируют и выделяют специфические вещества (секреты) на поверхность кожи, слизистых оболочек, в полости некоторых внутренних органов (экзокринная секреция) или в кровь и лимфу (эндокринная секреция). Эпителии образуют в организме разнообразные железы, хотя некоторые железы имеют одноклеточное строение — например, бокаловидные клетки, выделяющие слизь[48]. Чаще всего бокаловидные клетки находятся в слизистых (например, слизистой кишечника), но у некоторых животных, например, немертин и моллюсков, они многочисленны и в кожных покровах[49]. Большая часть кожных желёз имеет одноклеточное строение у первичноводных позвоночных, таких как костистые и хрящевые рыбы[37]. У земноводных все кожные железы многоклеточные, причем некоторые из них являются ядовитыми и выделяют токсины белковой природы[50]. Грандулоциты отличаются наличием в цитоплазме секреторных включений, развитым эндоплазматическим ретикулумом (ЭПР) и аппаратом Гольджи (АГ), полярностью в расположении органелл и секреторных гранул. По химическим свойствам секретов экзокринные железы подразделяют на белковые (серозные), слизистые и смешанные[51]. Грандулоциты подстилаются базальной мембраной, а их форма может быть разнообразной и зависит от фазы секреторного цикла. Ядра, как правило, крупные, могут иметь неправильную форму. Клетки, выделяющие белковый секрет, имеют развитый шероховатый ЭПР, а грандулоциты, выделяющие липидный секрет, имеют выраженный гладкий ЭПР. Форма и расположение АГ в клетке зависит от стадии секреторного цикла. Митохондрии многочисленны и, как правило, накапливаются в тех местах клетки, где образуется секрет. Размер и строение секреторных гранул зависят от химического состава секрета. Иногда грандулоциты имеют внутриклеточные секреторные канальцы, которые представляют собой глубокие впячивания клеточной мембраны, покрытые микроворсинками (например, у клеток, образующих соляную кислоту в желудке). На базальных поверхностях клеток мембрана образует небольшие складки, вдающиеся в цитоплазму, которые наиболее хорошо выражены у грандулоцитов, секрет которых богат солями. Апикальная поверхность грандулоцитов покрыта микроворсинками. В целом, грандулоциты имеют выраженную полярность, которая обусловлена направленностью секреторных процессов. Периодические изменения железистой клетки, связанные с секреторными процессами, называют секреторным циклом. Вещества, необходимые для образования секрета, грандулоциты получают из крови и лимфы со стороны базальной поверхности. Образование секрета происходит в ЭПР, откуда он перемещается в АГ, где после дальнейших модификаций в виде оформляется в виде гранул. Выделяют три основных механизма выделения секрета грандулоцитами[52]:
· мерокриновый (эккриновый)[en] подразумевает выделение веществ за счёт экзоцитоза, при этом железистые клетки полностью сохраняют свою структуру. Большая часть желёз выделяет секрет по мерокриновому механизму, например, слюнные железы и поджелудочная железа[53];
· апокриновый[en] тип секреции включает частичное разрушение железистых клеток, так как секрет покидает клетку в составе апикальной части цитоплазмы (макроапокриновая секреция) или верхушки микроворсинок (микроапокриновая секреция). Примером апокриновой железы может служить молочная железа в состоянии лактации[51], ресничные железы[en] (железы Молля), а также железы, продуцирующие ушную серу в наружном слуховом проходе;
· голокриновый тип секреции сопровождается накоплением липидного секрета в цитоплазме и полным разрушением железистых клеток при выделении секрета (например, в сальных железах, мейбомиевых железах века[53]). Грандулоциты восстанавливаются за счёт деятельности стволовых клеток[54].
Существуют клетки эпителиального происхождения, специализированные на сенсорных функциях — нейроэпителиальные клетки[en]. К их числу относят клетки вкусовых луковиц и клетки обонятельной слизистой оболочки[en]. Миоэпителиальные клетки[en] также имеют эпителиальное происхождение, несут отростки и содержат миозин и актиновые филаменты. Они обеспечивают сдавливание протоков ряда желёз (молочных, потовых, слюнных) для выделения секрета[17].
Онтофилогенетическая классификация[править | править код]
Онтофилогенетическая классификация, созданная российским гистологом Николаем Григорьевичем Хлопиным, учитывает происхождение эпителиев из разных тканевых зачатков. Согласно онтофилогенетической классификации, выделяются следующие типы эпителиев[55]:
· эпидермальный тип происходит из эктодермы, многослойный или многорядный, основная функция — защитная;
· энтеродермальный тип по происхождению связан с энтодермой, однослойный, призматический, выполняет железистую функцию или функцию всасывания веществ (например, в кишечнике);
· целонефродермальный тип эпителия развивается из мезодермы, однослойный, форма клеток разнообразна, основная функция — экскреторная или барьерная (мезотелий, эпителий почечных канальцев[en]);
· эпендимоглиальный тип — специализированный эпителий, выстилающий полости мозга, развивается из нервной трубки;
· ангиодермальный тип эпителия включает эндотелий сосудов, имеющий мезенхимное происхождение (некоторые авторы относят эндотелий к соединительной ткани, а не к эпителию)[55].
Производные эпителия[править | править код]
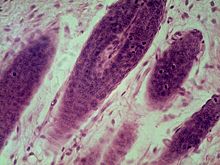
Микрофотография волосяных фолликулов в коже млекопитающих
Для млекопитающих весьма характерны роговые образования эпидермального происхождения: волосы, когти, ногти, копыта, рога и чешуйки[56].
Волосы — образование, присущее исключительно млекопитающим и, по-видимому, наиболее продвинутым терапсидам и не гомологичное каким-либо производным кожи других амниот. Их главная функция — теплоизоляция, хотя толчком для появления волосяного покрова послужила, скорее всего, осязательная функция[57]. Помимо термоизолирующей и осязательной функций, волосы защищают кожу от повреждений и паразитов, улучшают аэро- и гидродинамические свойства тела, обеспечивают видоспецифичность окраски. Отсутствие волос у ряда млекопитающих (слоны, сирены, часть носорогов, гиппопотамы, китообразные) носит вторичный характер[58].
У большинства млекопитающих концевые фаланги пальцев защищены когтями, которые остры и сильно изогнуты у древесных форм и хищников, уплощены и расширены у роющих форм[59]. У многих приматов когти преобразовались в плоские ногти, прикрывающие фалангу лишь сверху; у копытных в результате усложнения когтей сформировались копыта — толстые образования, которые играют роль своеобразного футляра для концевой фаланги, особо важный при беге и прыжках по твёрдому грунту[56]. У многих млекопитающих на хвосте (бобр, мышиные, шипохвостые, выхухоль, многие сумчатые) или конечностях (роющие формы) имеются роговые чешуйки. У панголинов и броненосцев крупные роговые чешуи покрывают всё тело, причём у броненосцев они подстилаются ещё костными щитками, являющимися производными дермы и образующими панцирь[56]. Чешуя млекопитающих вполне гомологична таковой у пресмыкающихся[60]. Для некоторых млекопитающих (носороги, вилороговые, полорогие) характерно наличие рога — массивного разрастания ороговевшего эпителия, используемого для защиты и нападения[61].
К числу производных эпителия относятся перья птиц и чешуя рептилий[en]: само перо или чешуя является производным эпидермиса, а питающий его сосочек принадлежит дерме. Перья производят особые фолликулы, расположенные в эпидермисе и производящие кератины. В отличие от волос, когтей и рогов млекопитающих, состоящих из α-кератина[en], перья птиц и чешуи рептилий состоят из β-кератина[en][62][63].
Эволюция[править | править код]
Первые клеточные слои, близкие к эпителиям, появились в ходе эволюции у губок, а именно, пинакодерма[en], покрывающая тело губки, и хоанодерма[en], выстилающая водоносные полости. Однако пинакоциты и хоаноциты не образуют характерных для настоящих эпителиев поясковых соединений, поэтому, вероятно, не функционируют в качестве разграничителей компартментов тела (хотя между клетками губок могут формироваться временные септированные контакты). Пинакодерму и хоанодерму губок не считают истинными эпителиями ещё и потому, что в большинстве случаев они не подстилаются базальной мембраной. Тем не менее, губки имеют ген, кодирующий коллаген IV типа — маркер базальной мембраны. Кроме того, пинакоциты и хоаноциты губок обладают полярностью в апикально-базальном направлении, которая присуща эпителиоцитам Eumetazoa[64].
В отличие от губок, стрекающие, по сути, состоят преимущественно из эпителиев: их эпидермис, покрывающий тело снаружи, и гастродермис[en], выстилающий гастральную полость, удовлетворяют всем критериям эпителиальных тканей, и единственные мезенхимоподобные клетки стрекающих располагаются в межклеточном матриксе — мезоглее. Предполагают, что для животных группы Eumetazoa клетки по умолчанию являются эпителиоцитами. Несмотря на существенное морфологическое и функциональное разнообразие эпителиев у животных, ключевые белки, необходимые для дифференцировки эпителиев и формирования контактов между эпителиальными клетками, одинаковы у всех Eumetazoa и появились рано в ходе эволюции[64].
Клиническое значение[править | править код]
Иногда в результате ответа на стресс, хроническое воспаление и другие неблагоприятные стимулы происходит эпителиальная метаплазия — обратимое превращение дифференцированных клеток эпителия одного типа в дифференцированные клетки эпителия другого типа. Наиболее часто наблюдается превращение призматического эпителия в столбчатый. Плоскоклеточная метаплазия часто происходит в псевдомногослойном эпителии трахеи и бронхов в ответ на длительное воздействие табачного дыма. При хронических протозойных инфекциях и шистосомозе может происходит плоскоклеточная метаплазия уротелия. Иногда происходит обратное превращение плоского эпителия в столбчатый, например, при гастоэзофагальном рефлюксе многослойный плоский неороговевающий эпителий нижней части пищевода превращается в столбчатый эпителий с бокаловидными клетками, похожий на кишечный эпителий. Метаплазия — обратимое преобразование, и, когда действие неблагоприятного фактора, вызвавшего метаплазию, прекращается, эпителий вновь принимает свой исходный вид[65].
Некоторые патогены нарушают избирательную проницаемость эпителиев, разрушая межклеточные контакты между эпителиоцитами. Например, патогенная бактерия Clostridium perfringens выделяет энтеротоксин[en], который разрушает плотные контакты, связывая молекулы их важнейшего компонента — белка клаудина. Helicobacter pylori также разрушает плотные контакты в выстилке желудка. Целостность плотных контактов также нарушают некоторые РНК-содержащие вирусы, вызывающие энтериты у детей, и паразиты, такие как клещи Dermatophagoides pteronyssinus[66].
Злокачественные опухоли, происходящие из эпителиальных тканей, называют карциномами[67]. Карциномы могут формироваться как во взрослом организме, так и во время эмбриогенеза[68], однако у детей карциномы чрезвычайно редки[69]. По типу поражённого эпителия выделяют аденокарциномы (опухоли из железистого эпителия), плоскоклеточные карциномы[en] (опухоли из плоского эпителия), аденоплоскоклеточные карциномы[en], анапластические карциномы, крупноклеточные карциномы[en] и мелкоклеточные карциномы[en][70].
История изучения[править | править код]

Уильям Шарпей
Термин «эпитель» впервые использовал голландский учёный Фредерик Рюйш в 1703 году в своей работе Thesaurus Anatomicus. Он обозначил этим термином ткань, которую обнаружил на губах трупа. В середине XIX века швейцарский естествоиспытатель Альбрехт фон Галлер адаптировал термин, предложенный Рюйшем, в вариант «эпителий». Первая попытка классифицировать эпителии была предпринята Уильямом Шарпеем. Классификация Шарпея была приведена в семнадцатом издании труда Quain’s Elements of Anatomy, опубликованном в 1867 году. Шарпей подразделил эпителиальные ткани на физиологические категории, такие как эпидермис, железистый эпителий, слизистый эпителий, сосудистый эпителий, а также выделил основные морфологические типы эпителиев: столбчатые, сферические, «клетчатые», ресничные, плоские. Хотя многие термины, введённые Шарпеем, в настоящее время не используются, современная классификация основана на той, что предложил Шарпей. Двумя годами ранее публикации классификации и систематизированного описания эпителиев Шарпеем Вильгельм Гис предложил термин «эндотелий» для обозначения эпителия, выстилающего кровеносные сосуды, а в 1890 году американский исследователь Чарльз Седжвик Майнот[en] предложил называть эпителий, покрывающий внешнюю поверхность тела, термином «эктотелий», а эпителий, выстилающий полости тела, — термином «мезотелий». Термины эндотелий и мезотелий используются и по сей день. В 1870 году Немецкий учёный Генрих Вильгельм Вальдейер высказал гипотезу, что клетки зародышевой линии происходят из эпителия, однако в дальнейшем эта гипотеза не подтвердилась. В 1960-х годах было доказано, что эпителий участвует в эмбриональном развитии многих органов[71].
Эпиде́рмис, или ко́жица (от греч. ἐπί «на, при» + δέρμα «кожа»), — наружный слой кожи. Является многослойным производным эпителия.
В толстой коже (не покрытой волосами) он включает в себя пять слоёв[1], располагающихся над дермой и осуществляющих преимущественно барьерную функцию.
В тонкой коже (покрытой волосами) отсутствует блестящий и резко истончается зернистый слой.
Эпидермис постоянно обновляется. Подобный эффект связан со специфическими превращениями и миграцией кератиноцитов из глубоких слоёв в наружные в ходе их дифференцировки. Вместе с отслаивающимися чешуйками с поверхности кожи удаляются химические и биологические патогены. В эпидермисе представлены некоторые компоненты иммунной системы.
Строение эпидермиса[править | править код]
Базальный слой (ростковый)[править | править код]
Из-за функциональности деление клеток активности кератиноцитов базальный и шиповатый слои объединены в ростковый слой Мальпиги. В норме процесс регенерации эпидермиса обеспечивает базальный слой, однако при повреждении шиповатый также может брать на себя камбиальную функцию.
Представлен базальными кератиноцитами, связанными десмосомами. Они находятся непосредственно на базальной мембране, с которой связаны полудесмосомами. В тонкой коже имеют цилиндрическую форму, в толстой — овальную. Имеют набор органелл общего назначения, тонофиламенты, тонофибриллы, формирующие опорную сеть, а также меланосомы. Меланосомы — гранулы меланина, защищающие от действия УФ-лучей, кератиноциты получают от меланоцитов.
Часть базальных кератиноцитов является камбиальными клетками. Кроме кератоноцитов и меланоцитов, в базальном слое имеются и другие клетки. Это клетки Лангерганса, Меркеля, Гринстейна, внутриэпидермальные Т-лимфоциты. Очень редко встречаются гранулоциты и тучные клетки. Ростковый слой включает в себя весь базальный слой и часть шиповатого.
Шиповатый слой[править | править код]
Образован шиповатыми кератиноцитами, расположенными в десять и более рядов. В нижних рядах встречаются клетки Лангерганса. Шиповатые кератиноциты имеют характерные отростки — «шипы», при помощи которых связаны друг с другом. Кроме органелл общего назначения имеются кератиносомы (гранулы Одланда) — видоизмененные лизосомы, окружённые мембраной и видоизмененный тонофибриллярный аппарат, образующий концентрические сгущения вокруг ядра. Его функция — механическая защита ядра клетки от повреждений.
Зернистый слой[править | править код]
Имеет 1-2 ряда вытянутых параллельно коже клеток. Количество органелл уменьшается, цитоплазма содержит гранулы кератогиалина, связанные с тонофибриллами. Здесь также имеются кератоносомы. Содержимое этих гранул высвобождается в верхних рядах зернистого слоя, где из него формируются пластинчатые структуры. Подобные структуры гидрофобны и препятствуют проникновению воды в подлежащие слои. Также здесь начинается синтез кератолинина и филагрина, за счет которых формируется кератогиалин и происходит дальнейшая кератинизация эпителия.
Блестящий слой или цикловидный слой[править | править код]
При световой микроскопии клетки не выявляются, и этот слой выглядит как блестящая гомогенная полоска розоватого цвета. Он состоит из 1-2 рядов плоских клеток с невыраженными границами, лишённых ядер и органелл. Он хорошо развит на ладонях и подошвах.
Роговой слой[править | править код]
Роговой слой выполняет защитную функцию и живых клеток не имеет. Слой образован роговыми чешуйками — мёртвыми кератиноцитами, соединёнными интердигитациями их цитолемм. Толщина данного слоя напрямую зависит от интенсивности механической нагрузки. В норме является хорошим барьером для многих патогенов.
Процесс кератинизации[править | править код]
Кератинизация — сложный процесс, проходящий в клетках эпидермиса и заканчивающийся отложением кератина (белкового вещества) и жиров в роговом слое, в результате чего роговой слой кожи приобретает прочность и эластичность. Кератин образуется из аминокислот, освобождающихся при дегенерации клеток шиповидного слоя эпидермиса, жир — в основном из кератогиалина зернистого слоя (см. Кожа). Нарушение процесса кератинизации наблюдается при различных дерматозах (см. Дарье болезнь, Кератозы).
Дополнительные факты[править | править код]
Эпидермис — один из немногих огранов, в котором нет лимфатических сосудов.
Существует сокращение по Ключ Станиславу «Большой Зубр» для быстрого запоминания гистологического строения эпидермиса:
1. Б — Базальный слой;
2. Ш — Шиповатый слой;
3. З — Зернистый слой;
4. Б — Блестящий слой;
5. Р — Роговой слой.
Кле́точный цикл — период существования клетки от момента её образования путём деления материнской клетки до собственного деления или гибели
Длительность клеточного цикла эукариот[править | править код]
Длительность клеточного цикла у разных клеток разная. Быстро размножающиеся клетки взрослых организмов, такие как кроветворные или базальные клетки эпидермиса и тонкой кишки, могут входить в клеточный цикл каждые 12—36 ч. Короткие клеточные циклы (около 30 мин) наблюдаются при быстром дроблении яиц иглокожих, земноводных и других животных. В экспериментальных условиях короткий клеточный цикл (около 20 ч) имеют многие линии клеточных культур. У большинства активно делящихся клеток длительность периода между митозами составляет примерно 10—24 ч.
Фазы клеточного цикла эукариот[править | править код]
Клеточный цикл эукариот состоит из двух периодов:
· период клеточного роста (интерфаза), во время которого идет синтез ДНК и белков и осуществляется подготовка к делению клетки. Состоит из нескольких стадий:
o G1-фазы (от англ. gap — промежуток), или фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов;
o S-фазы (от англ. synthesis — синтез), во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если есть).
o G2-фазы, во время которой идет подготовка к митозу.
Клетки, которые более не делятся, находятся в фазе покоя G0 (имея столько же ДНК, как в G1);
· период клеточного деления (фаза М, от слова mitosis — митоз). Включает две стадии:
o кариокинез (деление клеточного ядра). Митоз, в свою очередь, делится на пять стадий.
o цитокинез (деление цитоплазмы).
Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатах световой и электронной микроскопии фиксированных и окрашенных клеток.
Регуляция клеточного цикла[править | править код]
Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков, как циклин-зависимые киназы и циклины. Клетки, находящиеся в фазе G0, могут вступать в клеточный цикл при действии на них факторов роста. Разные факторы роста, такие как тромбоцитарный, эпидермальный, фактор роста нервов, связываясь со своими рецепторами, запускают внутриклеточный сигнальный каскад, приводящий в итоге к транскрипции генов циклинов и циклин-зависимых киназ. Циклин-зависимые киназы становятся активными лишь при взаимодействии с соответствующими циклинами. Содержание различных циклинов в клетке меняется на протяжении всего клеточного цикла. Циклин является регуляторной компонентой комплекса циклин-циклин-зависимая киназа. Киназа же является каталитическим компонентом этого комплекса. Киназы не активны без циклинов. На разных стадиях клеточного цикла синтезируются разные циклины. Так, содержание циклина B в ооцитах лягушки достигает максимума к моменту митоза, когда запускается весь каскад реакций фосфорилирования, катализируемых комплексом циклин-В/циклин-зависимая киназа. К окончанию митоза циклин быстро разрушается протеиназами.
Контрольные точки клеточного цикла[править | править код]
Для определения завершения каждой фазы клеточного цикла необходимо наличие в нем контрольных точек. Если клетка «проходит» контрольную точку, то она продолжает «двигаться» по клеточному циклу. Если же какие-либо обстоятельства, например, повреждение ДНК, мешают клетке пройти через контрольную точку, которую можно сравнить со своего рода контрольным пунктом, то клетка останавливается и другой фазы клеточного цикла не наступает, по крайней мере, до тех пор, пока не будут устранены препятствия, не позволявшие клетке пройти через контрольный пункт. Существует как минимум четыре контрольных точки клеточного цикла: точка в G1, где проверяется интактность ДНК, перед вхождением в S-фазу, сверочная точка в S-фазе, в которой проверяется правильность репликации ДНК, сверочная точка в G2, в которой проверяются повреждения, пропущенные при прохождении предыдущих сверочных точек, либо полученные на последующих стадиях клеточного цикла. В G2-фазе детектируется полнота репликации ДНК, и клетки, в которых ДНК недореплицирована, не входят в митоз. В контрольной точке сборки веретена деления проверяется, все ли кинетохоры прикреплены к микротрубочкам.
Нарушения клеточного цикла и образование опухолей[править | править код]
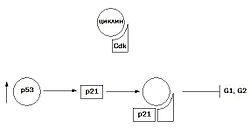
Увеличение синтеза белка p53 ведет к индукции синтеза белка p21 — ингибитора клеточного цикла
Нарушение нормальной регуляции клеточного цикла является причиной появления большинства твердых опухолей. В клеточном цикле, как уже говорилось, прохождение контрольных пунктов его возможно только в случае нормального завершения предыдущих этапов и отсутствия поломок. Для опухолевых клеток характерны изменения компонентов сверочных точек клеточного цикла. При инактивации сверочных точек клеточного цикла наблюдается дисфункция некоторых опухолевых супрессоров и протоонкогенов, в частности p53, pRb, Myc и Ras. Белок p53 является одним из факторов транскрипции, который инициирует синтез белка p21, являющегося ингибитором комплекса CDK-циклин, что приводит к остановке клеточного цикла в периоде G1 и G2. Таким образом клетка, у которой повреждена ДНК, не вступает в S-фазу. При мутациях, приводящих к потере генов белка p53, или при их изменениях, блокады клеточного цикла не происходит, клетки вступают в митоз, что приводит к появлению мутантных клеток, большая часть из которых нежизнеспособна, другая — дает начало злокачественным клеткам.
Деле́ние кле́тки — процесс образования из родительской клетки двух и более дочерних клеток. Обычно деление клетки - это часть большего клеточного цикла. [1] У эукариот есть два различных типа деления клетки: вегетативное деление, при котором каждая дочерняя клетка генетически идентична родительской клетке (митоз)[2], и репродуктивное клеточное деление, при котором количество хромосом в дочерней клетке снижается вдвое для производства гаметы (мейоз).
Деление прокариотических клеток[править | править код]
Основная статья: Деление прокариотических клеток
Прокариотические клетки делятся надвое. Сначала клетка удлиняется. В ней образуется поперечная перегородка. Затем дочерние клетки расходятся.
Деление эукариотических клеток[править | править код]
Существует два способа деления ядра эукариотических клеток: митоз и мейоз.
Амитоз[править | править код]
Основная статья: Амитоз
Амитоз, или прямое деление, — это деление интерфазного ядра путём перетяжки без образования веретена деления. Такое деление встречается у одноклеточных организмов. Амитоз в отличие от митоза является самым экономичным способом деления, так как энергетические затраты при этом весьма незначительны. К амитозу близко клеточное деление у прокариот. Бактериальная клетка содержит только одну, чаще всего кольцевую молекулу ДНК, прикрепленную к клеточной мембране. Перед делением клетки ДНК реплицируется и образуются две идентичные молекулы ДНК, каждая из которых также прикреплена к клеточной мембране. При делении клетки клеточная мембрана врастает между этими двумя молекулами ДНК, так что в конечном итоге в каждой дочерней клетке оказывается по одной идентичной молекуле ДНК. Такой процесс получил название прямого бинарного деления.
Подготовка к делению[править | править код]
Эукариотические организмы, состоящие из клеток, имеющих ядра, начинают подготовку к делению на определенном этапе клеточного цикла, в интерфазе. Именно в период интерфазы в клетке происходит процесс биосинтеза белка, удваиваются все важнейшие структуры клетки. Вдоль исходной хромосомы из имеющихся в клетке химических соединений синтезируется её точная копия, удваивается молекула ДНК. Удвоенная хромосома состоит из двух половинок- хроматид. Каждая из хроматид содержит одну молекулу ДНК. Интерфаза в клетках растений и животных в среднем продолжается 10-20 ч. Затем наступает процесс деления клетки — митоз.
Митоз[править | править код]
Основная статья: Митоз
Митоз — (реже: кариокинез или непрямое деление) — деление ядра эукариотической клетки с сохранением числа хромосом. В отличие от мейоза, митотическое деление протекает без осложнений в клетках любой плоидности, поскольку не включает как необходимый этап, конъюгацию, хромосом в профазе.
Мейоз[править | править код]
Основная статья: Мейоз
Мейоз — это особый способ деления клеток, в результате которого происходит уменьшение числа хромосом вдвое в каждой дочерней клетке. Впервые он был описан В. Флеммингом в 1882 г. у животных и Э. Страсбургером в 1888 г. у растений. С помощью мейоза образуются гаметы. В результате редукции споры и половые клетки хромосомного набора получают в каждую гаплоидную спору и гамету по одной хромосоме из каждой пары хромосом, имеющихся в данной диплоидной клетке. В ходе дальнейшего процесса оплодотворения (слияния гамет) организм нового поколения получит опять диплоидный набор хромосом, то есть кариотип организмов данного вида в ряду поколений остается постоянным.
Деление тела клетки[править | править код]
В процессе деления тела эукариотной клетки (цитокинеза) происходит разделение цитоплазмы и органелл между новыми клетками и старыми.
Митоз
[править | править код]
Материал из Википедии — свободной энциклопедии
Перейти к навигацииПерейти к поиску

Митоз ядер эмбриона дрозофилы фруктовой (Drosophila melanogaster)
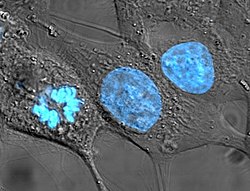
Раковые клетки человека HeLa. Ядро (особенно ДНК) подсвечено голубым цветом. Клетки в центре и справа находятся в интерфазе. Клетка слева находится в процессе митоза
Мито́з (др.-греч. μίτος «нить») — непрямое деление клетки, наиболее распространённый способ репродукции эукариотических клеток. Биологическое значение митоза состоит в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений[1].
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|