- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Теории слуха
Глава 6
Анализаторы[I]
6.1. ОБЩАЯ ХАРАКТЕРИСТИКА АНАЛИЗАТОРОВ
Живой организм постоянно воспринимает воздействия внешней и внутренней среды. Совокупность возбудимых структур периферической и центральной нервной системы, осуществляющих восприятие и анализ воздействий окружающей и внутренней среды, получила название анализаторов.
Структурно-функциональная организация анализаторов.Все структуры, входящие в состав анализаторов, относятся к афферентным, т.е. проводящим возбуждения от периферии в ЦНС. Классические представления И. П. Павлова об анализаторе включают в его состав три части: периферический отдел, проводниковый отдел и центральный конец (рис. 6.1). Анализаторы разлагают внешний мир на элементы, а затем трансформируют раздражение в ощущение, после которого формируется определенная тактика поведения организма.
Клетки коры больших полушарий осуществляют, по И. П. Павлову, высший анализ и синтез воздействий внутренней и внешней среды.
Периферический отдел анализаторовначинается с рецепторов, хотя в некоторых анализаторах в этот отдел могут быть вклю-
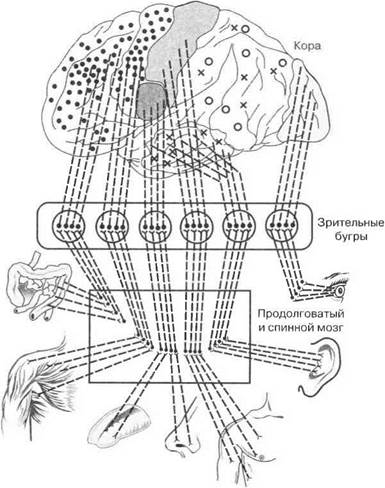
Рис. 6.1.Общая схема строения анализаторов
чены специальные органы чувств, которые помимо рецепторов включают специальные вспомогательные образования для наилучшего восприятия действующих раздражителей. Например, глаз как орган зрения помимо сетчатки, где расположены фоторецепторы, имеет сложное строение: глазное яблоко, его мышцы, веки, хрусталик и др.
Органы чувств — специализированные образования, разлагающие внешние воздействия на такие составляющие, которые могут быть восприняты соответствующими рецепторами.
Проводниковый отдел анализатороввключает не только нервные волокна, непосредственно отходящие от рецепторов, но и все афферентные нейроны, обеспечивающие первичный анализ и передачу возбуждений в центральный отдел анализатора. В потоках импульсов возбуждения, распространяющихся по проводящим путям, в их своеобразном рисунке — паттерне — закодирована специфическая информация о параметрах действующих раздражителей. Поскольку различные нервные волокна имеют различную скорость проведения возбуждения, это определяет опережение или запаздывание распространения в ЦНС отдельных возбуждений рецепторов, что также имеет информационный смысл. Анализ этой информации начинается как на уровне первичных афферентных нервных клеток, так и в последующих спинальных, стволовых и подкорковых структурах ЦНС.
Центральный отдел анализаторов.Различные проводящие афферентные пути через возбуждение специальных подкорковых структур в конечном счете приносят импульсы возбуждения в соответствующие области коры большого мозга — высшему центральному конечному звену любого анализатора. Вместе со специфическими афферентными возбуждениями в кору поступают и неспецифические восходящие возбуждения, которые формируются на уровне подкорковых активирующих структур головного мозга — ретикулярной формации, гипоталамуса и др. В свою очередь возбуждения из коры головного мозга адресуются лимбическим образованиям, формируя тем самым эмоциональное восприятие раздражения.
Кора головного мозга у человека включает четыре отдела: лобный, теменной, затылочный и височный, каждый из которых в свою очередь подразделяется на проекционные и ассоциативные области. Электрическое раздражение проекционных областей при нейрохирургических операциях у человека сопровождается специфическими ощущениями. Например, при раздражении соматосенсорной коры возникает чувство прикосновения к коже, при раздражении зрительной коры возникают зрительные образы, при раздражении слуховой коры появляется ощущение звучания. При раздражении ассоциативных областей — лобной, височной, теменно-затылочной — возникают интегральные образы, нередко связанные с прошлым жизненным опытом субъектов.
6.2. АНАЛИЗАТОРЫ В СИСТЕМНОЙ ОРГАНИЗАЦИИ ПОВЕДЕНИЯ
В системной организации внутримозговых процессов при формировании поведения афферентные потоки возбуждения всегда имеют конкретное функциональное значение не только по сенсорной модальности афферентного возбуждения, но и по их роли в механизмах системной архитектоники поведенческих актов, в частности в механизмах афферентного синтеза и аппарата предвидения результатов действия. В системной организации поведения различают следующие потоки афферентных возбуждений: обстановочную афферентацию, пусковую афферентацию и обратную афферентацию.
Обстановочная афферентация.Участвует в формировании стадии афферентного синтеза. Она складывается из воздействий на организм всей совокупности внешних факторов, составляющих конкретную обстановку, на фоне которой формируется поведенческий акт. По отношению к человеку обстановочными раздражителями выступают многочисленные социальные факторы: нравственные, культурные, бытовые и др.
Пусковая афферентация.Под пусковой афферентацией понимают афферентные возбуждения, возникающие при воздействии на организм стимулов, непосредственно развертывающих поведенческий акт. В качестве пусковых стимулов могут выступать многочисленные условные раздражители и время.
Обратная афферентация.Афферентные возбуждения, приносящие в структуры головного мозга информацию о возникшей в организме потребности и об успешности достижения результатов целенаправленного поведения. В функциональных системах организма обратная афферентация в виде нервной импульсации от интерорецепторов и гуморальных влияний на центры нервной системы постоянно сигнализирует о степени достижения полезных для организма результатов, в частности об оптимальном уровне тех или иных показателей внутренней среды организма. В любом поведенческом акте, который направлен на получение какого-либо приспособительного, т.е. полезного для организма, результата, обратная афферентация информирует об этом результате совершенного действия, давая возможность организму в целом оценить степень успеха выполняемого им действия и своевременно исправить ошибки. Например, у человека на основе метаболических процессов и взаимодействия организма и внешней среды может созреть намерение выпить стакан чая. Он протягивает руку к стакану с чаем, берет его. Тактильное возбуждение ладони поверхностью стакана, температурное, весовое, наконец, зрительное раздражение от контакта руки со стаканом — все эти афферентные раздражения в сумме дают обратную информацию о том, что результат действия соответствует исходному намерению. К тому же при осуществлении этого действия само продвижение руки к стакану непрерывно регулируется проприоцептивной сигнализацией, свидетельствующей о правильном и соответствующем распределении сокращенных мышц, о степени напряжения руки, о высоте ее положения по отношению к намеченной цели и т.д.
Направляющая и результативная обратная афферентация.Обратная афферентация, возникающая при двигательном акте, разделяется на две различные категории: направляющую движение и результативную афферентацию. Направляющая афферентация представлена в основном импульсами от проприорецепторов мышц, осуществляющих движение, результативная — всегда комплексная и может включать афферентные потоки от фоторецепторов сетчатки, тактильных, вкусовых, слуховых и прочих рецепторов. В свою очередь обратная результативная афферентация подразделяется на поэтапную обратную афферентацию и санкционирующую обратную афферентацию, которая возникает при достижении субъектом потребного результата, закрепляет наиболее успешную интеграцию афферентных возбуждений и завершает функциональный системок- вант поведения (например, «хочу пить» — «напился»).
Анализаторы в системной организации физиологических функций — динамические самонастраивающиеся и саморегулирующиеся организации.
Механизмы саморегуляции анализаторов.Восприятие действующих на организм раздражителей является активным процессом, в котором каждый анализатор представляет собой сложную самонастраивающуюся афферентно-эфферентную организацию с множеством обратных связей. Уже на уровне рецепторных клеток и
даже молекулярных процессов в рецепторах осуществляется отбор информации за счет местных механизмов саморегуляции активности рецепторов. Благодаря антидромному движению афферентных импульсов по разветвлениям чувствительных волокон осуществляется антидромный механизм, тормозящий активность окружающих рецепторов, находящихся как в пределах поля активируемого рецептора, так и в соседних перекрывающихся и неперекрывающихся рецептивных полях. Местный механизм саморегуляции рецепторов происходит без участия центральных нейронов и выполняет задачи ограничения афферентного потока и подчеркивания пространственно-временного контраста раздражителя.
Наряду с антидромным в саморегуляции анализаторов участвует и центробежный механизм контроля афферентации, который может иметь как тормозной, так и облегчающий характер и осуществляться с помощью различных эфферентных путей и специальных эфферентных синапсов. Антидромный и центробежный механизмы осуществляют контроль функциональной активности рецепторных клеток, обеспечивая избирательное реагирование организма на факторы внешней среды. В механизмах саморегуляции, чувствительности рецепторов ведущая роль принадлежит доминирующей мотивации (см. разд. 8.5).
Восприятие.Восприятие воздействия определяется, прежде всего, ощущением. Эмоциональные ощущения порождаются возбуждением лимбических структур головного мозга. Как указывалось в гл. 3, в лимбические структуры распространяются проекции практически всех афферентных путей, в частности зрительных, обонятельных, висцеральных, слуховых и болевых. Лимбические структуры мозга осуществляют эмоциональную информационную оценку действующих на организм внешних и внутренних раздражений.
Наряду с этим детальная оценка зрительных, слуховых, обонятельных, вкусовых, осязательных и болевых воздействий осуществляется в соответствующих проекционных зонах коры больших полушарий.
Количество чувств в настоящее время расширилось. К шести классическим чувствам (зрение, вкус, обоняние, слух, осязание, ощущение боли) прибавились чувства равновесия, ориентации в пространстве (homing), ощущение времени, ощущение препятствий на расстоянии (например, у слепых людей и у летучих мышей), эмоциональные ощущения голода, жажды, половых возбуждений, страха, агрессии, температурных мотиваций, позывов и т.д.
На уровне нейронов мозга испытанные субъектами ощущения фиксируются в виде эмоциональных стереотипов. На основе эмоциональных ощущений в процессе онтогенеза в мозговых структурах складываются:
• температурная схема тела (показатель рассогласования — чихание);
• осязательная схема тела (показатель рассогласования — зуд);
• схема положения тела в пространстве (показатель рассогласования — головокружение);
• схемы отпечатков музыкальных мелодий;
• электромагнитная схема тела;
• словесные схемы действительности и др.
Отмеченные информационные схемы мозга в значительной степени определяют поведение живых существ. Их нарушение у отдельных субъектов может вызвать невротические реакции.
Восприятие тесно связано со свойством мозга опережающее, на основе механизмов памяти отражать действительность. Аппарат акцептора результата действия позволяет живым существам постоянно оценивать биологическую значимость действующих на них раздражителей, их полезность или вредность.
Ведущая роль в процессах восприятия принадлежит доминирующим мотивациям (см. разд. 8.5), которые избирательно настраивают нейроны мозга, их рецепторы, а также периферические рецепторы на восприятие тех предметов внешнего мира, которые удовлетворяют исходные потребности организма. При этом каждый вид животных оценивает параметры достигнутых результатов своими ведущими анализаторами.
6.3. ЧАСТНАЯ ФИЗИОЛОГИЯ АНАЛИЗАТОРОВ
Поведение живых организмов в окружающей среде не является пассивным отражением воздействующих раздражителей. Живые организмы, как правило, настойчиво ищут потребные раздражители и активно к ним стремятся, взаимодействуя с одними из них и активно избегая других, избирательно настраивая по отношению к ним свои анализаторы.
Активное стремление субъектов к раздражителям внешней среды определяется, прежде всего, их исходными доминирующими потребностями и пропускной способностью к передаче информации соответствующих анализаторов. У человека наибольшей пропускной способностью обладает зрительный анализатор, который в единицу времени передает в ЦНС более 70 % информации; 25—28 % информации доставляет в ЦНС слуховой анализатор и 2—5 % информации — остальные анализаторы.
6.3.1. Зрительный анализатор
Светопреломляющие структуры глаза.Оптические светопреломляющие среды глаза фокусируют изображение на сетчатке. Основными светопреломляющими средами глаза, которые обеспечивают формирование на сетчатке реального, уменьшенного и перевернутого изображения объекта внешнего мира, являются роговица, радужная оболочка, хрусталик, камерная влага и стекловидное тело.
Радужная оболочка образует зрачок, который имеет у человека круглую форму и одинаковый диаметр в обоих глазах. Светопреломляющая способность хрусталика и диаметр зрачка изменяются при сокращении гладких мышц глаза. Зрачковая реакция на свет является механизмом снижения количества света, падающего на сетчатку при сильном освещении (сужение зрачка), или повышения количества света при слабом освещении за счет увеличения ширины зрачка. Сужение зрачка обеспечивается сокращением круговой мышцы — сфинктера радужной оболочки, иннервируемой парасимпатическими волокнами. Расширение зрачка обеспечивается сокращением мышцы-дилататора с радиальными волокнами в радужной оболочке, иннервируемой симпатическими волокнами.
Аккомодация — приспособление глаза к ясному видению объектов, находящихся на разных расстояниях. Главную роль в аккомодации играет хрусталик, изменяющий свою кривизну, благодаря чему изображение предметов фокусируется на сетчатке. Выпуклость хрусталика определяется сокращением ресничных мышц. С другой стороны, хрусталик заключен в капсулу, которую всегда растягивают (уплощают) волокна ресничного пояска (Циннова связка). Ресничные мышцы иннервируются парасимпатическими волокнами глазодвигательного нерва. Атропин блокирует передачу возбуждения к этим мышцам, что ограничивает аккомодацию глаза при рассматривании близких предметов.
Далекие предметы нормальный глаз рассматривает без напряжения аккомодации, т.е. без сокращения ресничной мышцы. Ближайшая точка ясного видения благодаря аккомодации находится на расстоянии 10 см от глаза. Точка ясного видения изменяется при различных эмоциональных состояниях человека.
Сетчатка глаза.Физиологические механизмы опознания зрительных объектов начинаются с первичной обработки зрительной информации в сетчатке глаза, которая является периферической рецепторной структурой зрительного анализатора. Сетчатка расположена на внутренней поверхности задней сферы глазного яблока и состоит из клеток пигментного эпителия, фоторецепторов и четырех слоев, образованных различными нервными клетками и клетками глии.
Фоторецепторы сетчатки.Основными зрительными рецепторами, расположенными в сетчатке, являются палочки и колбочки. Восприятие фоторецепторами световых волн лежит в диапазоне от 400 до 750 нм. У человека рецепторный слой сетчатки состоит из 120 млн палочек и 6 млн колбочек, которые имеют различное гистологическое строение. Центральная ямка сетчатки имеет наибольшую плотность колбочек, а периферия сетчатки содержит почти одни палочки. Колбочки воспринимают цвета и функционируют в условиях яркой освещенности объектов, в то время как палочки воспринимают световые потоки в условиях сумерек — черно-белый свет.
Палочки и колбочки обращены своими наружными светочувствительными сегментами к пигментному эпителию. Пигментный эпителий играет решающую роль в ресинтезе зрительного пигмента после его обесцвечивания. Внутренние сегменты содержат крупные ядра, митохондрии и аппарат Гольджи. Здесь осуществляется синтез белка и обновление мембран наружных сегментов каждые 2—3 недели. В пресинаптических образованиях зрительных рецепторов выделяется нейромедиатор — глутамат.
Особенности морфофункциональной организации сетчатки.Сетчатка представляет собой довольно сложную нейронную сеть с горизонтальными и вертикальными связями между фоторецепторами и клетками. Биполярные клетки сетчатки передают сигналы от фоторецепторов в слой ганглиозных клеток и к амакриновым клет
кам (вертикальная связь). Горизонтальные и амакриновые клетки участвуют в горизонтальной передаче сигналов между соседними фоторецепторами и ганглиозными клетками (рис. 6.2). Сложность структурной организации и наличие в сетчатке процессов конвергенции и дивергенции сигналов позволяют рассматривать ее как часть мозга, вынесенную наружу.
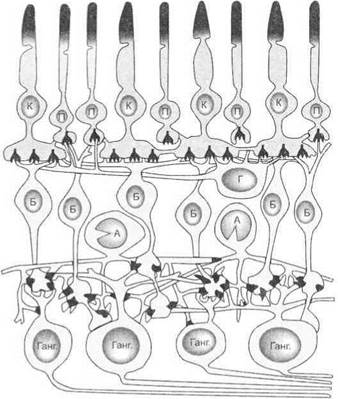
Рис. 6.2.Строение сетчатки:
К — колбочки; П — палочки; Б — биполяры; Г— горизонтальная клетка; А — амакриновая клетка; Ганг. — ганглиозные клетки
Зрительные пигменты. В палочках сетчатки содержится пигмент родопсин. В наружных сегментах колбочек содержится три типа зрительных пигментов к синей, зеленой и красной частям спектра цвета.
Красный колбочковый пигмент — йодопсин.
Зрительные пигменты состоят из белковой части (опсин) и хромоформной части (изомер 11-цис-ретиналь или альдегид витамина А). Источником ретинола являются каротиноиды. При недостатке в организме витамина А развивается «куриная слепота» — затруднение видимости в сумрачное время суток.
Молекулярные изменения при фоторецепции в палочках. При поглощении кванта света происходит изомеризация хромоформной части зрительных рецепторов: 11-цис-ретиналь превращается в транс-ретиналь. В результате происходят изменения в белковой опсиновой части рецепторов, родопсин обесцвечивается и переходит в метародопсин II. Метародопсин II взаимодействует с другим белком — трансдуцином и обменивает связанный с ним в темноте гуанозиндифосфат на гуанозинтрифосфат. Активированная молекула трансдуцина, в свою очередь, активирует одну молекулу фермента фосфодиэстеразы, которая разрушает молекулы цГМФ. В результате в цитоплазме наружного сегмента рецептора падает концентрация свободного цГМФ. Это, в свою очередь, приводит к закрытию ионных каналов в плазматической мембране наружного сегмента, которые были открыты в темноте и через которые внутрь рецепторных клеток входили ионы Na+ и Са2+. Уменьшение входа ионов Na+ приводит к гиперполяризации мембраны и возникновению рецепторного потенциала.
При действии света уменьшается выделение глутамата из пре- синаптических окончаний фоторецепторов, что приводит к гиперполяризации мембран ганглиозных клеток.
Цветовое зрение.В объяснении цветового зрения существует несколько теорий.
Трехкомпонентная теория М. В. Ломоносова — Г. Гельмгольца — К. Юнга. Согласно этой теории, цветовое восприятие обеспечивается зремя типами колбочек: к красному, зеленому и синему частям спектра цвета. При смешении этих основных цветов могут быть получены любые цвета.
Контрастная теория Э. Геринга строится на утверждении, что в колбочках содержатся три светочувствительных вещества — белочерное, красно-зеленое и желто-синее. Под влиянием одних световых лучей эти вещества распадаются, что вызывает ощущение белого, красного и желтого цветов. Другие световые лучи синтезируют исходные вещества и вызывают ощущение черного, зеленого и синего цветов.
Цветовая слепота.У определенных лиц, преимущественно у мужчин, отсутствуют гены в половой непарной Х-хромосоме. Вследствие этого они не воспринимают определенные цвета — дальтонизм (описал Д. Дальтон в XVIII веке).
Механизмы активности зрительных центров.При действии света в рецепторах и нейронах сетчатки генерируются электрические потенциалы.
Суммарный электрический ответ сетчатки — электроретино- грамма.
Электроретинограмма (ЭРГ). Интегративные функции сетчатки, связанные с первичным анализом световых раздражителей, находят отражение в изменениях ее суммарной биоэлектрической активности, регистрируемой в виде электроретинограммы (рис. 6.3).
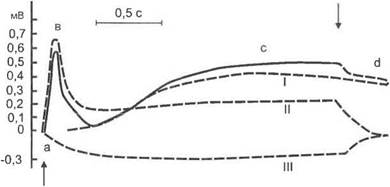
Рис. 6.3.Электроретинограмма:
а, Ь, с, d— основные волны электроретинограммы, образующиеся при алгебраическом сложении /, II и III компонентов; стрелками отмечены моменты включения и выключения света
Волна а отражает возбуждение внутренних сегментов фоторецепторов (поздний рецепторный потенциал) и горизонтальных клеток. Волна b возникает в результате активации глиальных клеток сетчатки ионами К+, выделяющимися при возбуждении биполярных и амакриновых нейронов. Волна с отражает активацию клеток пигментного эпителия, а волна d — горизонтальных клеток.
Рецептивные поля сетчатки. Конвергенция и дивергенция связей в сетчатке составляют основу организации рецептивных полей ее ганглиозных клеток. Рецептивное поле — область сетчатки, в пределах которой соответствующий раздражитель вызывает возбуждение или торможение данной ганглиозной клетки. Ганглиозная клетка генерирует потенциалы действия, которые по аксону распространяются в центральные отделы зрительного анализатора.
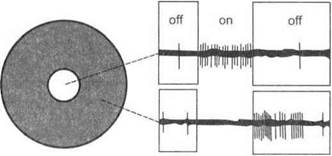
Ганглиозные нейроны реагируют на различные параметры световых воздействий. Описаны три типа ганглиозных клеток, реагирующих на включение (on-реакция) и выключение (off-реакция) света, а также на оба эти воздействия.
Диаметр рецептивных полей ганглиозных клеток в центре сетчатки значительно меньше, чем на периферии. Эти рецептивные поля имеют круглую форму: круглый возбудительный центр и концевая тормозящая периферическая зона и наоборот (рис. 6.4). Одно-
|
|
 | 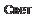 |
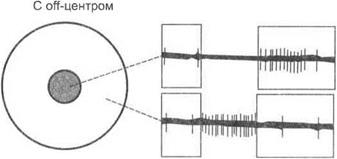
Рис. 6.4.Рецептивные поля ганглиозных клеток в сетчатке
временные возбуждения двух близко расположенных ганглиозных клеток приводят к их взаимному торможению. В основе этого явления лежит латеральное торможение. Благодаря круглой форме рецептивные поля ганглиозных клеток сетчатки производят так называемое поточечное мозаичное отражение светового изображения.
Зрительные пути.Схема зрительных путей в головном мозге человека представлена на рис. 6.5.
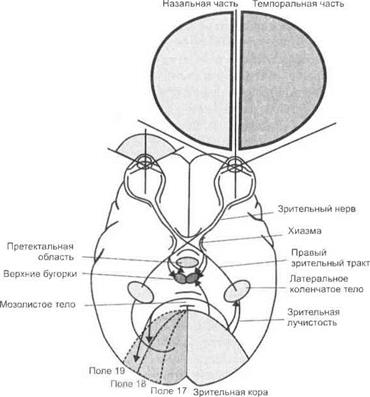
Рис. 6.5.Зрительные пути в головном мозге человека (схема)
Аксоны ганглиозных клеток дают начало зрительному нерву. Зрительный нерв состоит из 106 аксонов, которые передают сигналы от 13 х 107 фоторецепторов сетчатки одного глаза. Правый и левый зрительные нервы сливаются у основания черепа, образуя перекрест, где нервные волокна, идущие от внутренних (носовых) половин обеих сетчаток, пересекаются и переходят на противоположную сторону. Волокна, идущие от наружных (височных) половин каждой сетчатки, продолжают идти с той же стороны, объединяясь вместе с перекрещенным пучком аксонов из контралатерального зрительного нерва, образуя зрительный тракт. Зрительный тракт заканчивается в первичных подкорковых центрах зрительного анализатора, к которым относятся латеральные коленчатые тела, верхние бугорки четверохолмия и претектальная область ствола мозга.
Латеральные коленчатые тела.Латеральные коленчатые тела являются первой структурой ЦНС, где происходит переключение импульсов возбуждения на пути между сетчаткой и корой головного мозга. Нейроны латерального коленчатого тела обладают простыми рецептивными полями подобно ганглиозным клеткам сетчатки. Нейроны сетчатки и латерального коленчатого тела производят анализ зрительных стимулов, оценивая их цветовые характеристики, пространственный контраст и среднюю освещенность в различных участках поля зрения. В латеральных коленчатых телах начинается бинокулярное взаимодействие от сетчатки правого и левого глаза. Взаимодействие возбуждений на нейронах латерального коленчатого тела осуществляется на основе механизма реципрокного торможения.
Рецептивные поля нейронов латерального коленчатого тела тоже круглые, но меньшего размера, чем в сетчатке.
Здесь происходит взаимодействие афферентных сигналов, приходящих от сетчатки с эфферентными возбуждениями, формируемыми в различных отделах ЦНС, что обеспечивает процессы избирательного зрительного внимания.
Верхние бугорки четверохолмия.Нейроны верхних бугорков четверохолмия преимущественно реагируют на движущиеся световые стимулы. Некоторые из них возбуждаются только в том случае, когда стимул движется через их рецептивное поле в определенном направлении. В более глубоких слоях бугорков находятся нервные клетки, которые возбуждаются в моменты, предшествующие движениям глазных яблок. Эти клетки включены также в механизмы управления целенаправленным движением глаз.
Корковые центры зрительного анализатора.Аксоны нейронов латерального коленчатого тела расходятся в виде лучей (зрительная радиация) и оканчиваются, в основном, в поле 17 затылочной доли коры головного мозга (первичная зрительная кора). Первичная зрительная кора образует связи с полями 18 и 19 (вторичная и третичная зрительная кора), а также с верхними бугорками четверохолмия. В первичной, вторичной и третичной зрительной коре представлена контралатеральная половина поля зрения.
Нейроны зрительной коры имеют не круглые, а вытянутые по горизонтали и вертикали или в одном из косых направлений рецептивные поля. Благодаря этому они выделяют из цельного изображения отдельные фрагменты линий с различной ориентацией. В каждом участке зрительной коры сконцентрированы нейроны с одинаковой ориентацией зрительных раздражений. Они образуют колонки, локализующиеся вертикально через все слои коры.
Обработка возбуждений нейронами зрительной коры.Метод микроэлектродной регистрации электрической активности отдельных нейронов у высших млекопитающих позволил подробно изучить особенности обработки зрительных возбуждений нейронами зрительной коры. По сравнению с нейронами латерального коленчатого тела корковые зрительные нейроны осуществляют более сложный анализ световых стимулов. Специализация нейронов зрительной коры определяется свойствами их рецептивных полей, которые подразделяются на простые, сложные и сверхсложные.
Зрительные корковые нейроны с простым рецептивным полем активируются при воздействии на фоторецепторы световым стимулом в виде полосы, определенным образом расположенной в пространстве (вертикально, горизонтально или под углом) (рис. 6.6). Для активации нейронов со сложным рецептивным полем необходимы не только оформленный и пространственно-ориентированный стимул, но и определенное направление его движения. Например, оптимально пространственно оформленный стимул оказывает возбуждающий эффект только в том случае, если его отражение движется по возбудительному рецептивному полю слева направо, а движение в обратном направлении не активируют нейроны. Чувствительность корковых зрительных нейронов к движению и направлению внешних предметов обусловлена тем, что изображение неподвижного предмета всегда смещается по сетчатке из-за непрерывного движения глаз и тела. Нейроны со сложными рецептивными полями, как правило, имеют их для каждого глаза и могут

|
Нейроны со сверхсложными полями находятся в основном в полях 18 и 19 коры большого мозга. Они возбуждаются только в том случае, если на их возбуждающее рецептивное поле воздействует световой стимул с большим количеством зрительных параметров (форма, пространственная ориентация, направление движения, значения углов на границе, свет и темнота и др.).
Движение глаз и бинокулярное зрение.Перемещение зрительного изображения по сетчатке достигается постоянными сканирующими движениями глаз — саккадами, что является необходимым условием зрительного восприятия видимых предметов. Периоды фиксации длятся в среднем от 0,15 до 2 с. Сканирующие движения глаз крайне необходимы, например, при чтении текста. Они контролируются нейронами со сложными рецептивными полями 18 и 19 зрительной коры головного мозга. Смена коротких периодов фиксации на сетчатке изображений объекта за счет движений глаз создает в головном мозге стационарное «изображение» — субъективный образ внешнего мира.
Саккадические движения глаз, как и любая двигательная активность человека, могут служить показателями его внутреннего психологического состояния. Такие состояния человека, как внимание, интерес, усталость, страх, существенно влияют на характеристики движений глаз, отражая общее эмоциональное состояние.
Глазные мышцы и формы движения глаз.Глаз человека приводится в движение шестью наружными мышцами. Эти мышцы создают горизонтальные, вертикальные и вращательные движения глазного яблока, обеспечивая перемещение по сетчатке сфокусированного изображения светового стимула. Движения двух глаз могут быть содружественными, т.е. двигаться вместе вверх, вниз, влево или вправо, а могут быть конвергентными (сходящимися) или дивергентными (расходящимися) при рассматривании близко или далеко расположенного предмета. Конвергентные и дивергентные движения глаз обеспечивают восприятие глубины пространства.
Бинокулярное зрение.Механизм регуляции одновременного движения правого и левого глазных яблок создает эффект бинокулярного зрения. При этом конвергентные и дивергентные движения обоих глаз управляются нейронами, находящимися как в подкорковых структурах, так и в коре головного мозга. Центры бинокулярного зрения находятся в области ретикулярной формации среднего мозга, в верхних бугорках четверохолмия и в претектальной области. Ретикулярная формация среднего мозга является интегрирующим центром, получающим информацию по афферентным путям не только от верхних бугорков четверохолмия и зрительных корковых полей 18 и 19, но и непосредственно от фоторецепторов сетчатки. Ядра глазодвигательных нервов находятся также под корригирующим влиянием мозжечка, обеспечивающим более точное сведение зрительных осей обоих глаз к одной и той же точке. В мозжечке вестибулярные и зрительные сигналы взаимодействуют с сигналами, отражающими положение головы и глаз. Интегрированные возбуждения передаются по мозжечковым эфферентным путям к стволовым центрам, управляющим бинокулярным зрением.
6.3.2. Слуховой анализатор
Максимальная чувствительность слуха человека лежит в области частот от 1000 до 4000 Гц. Шепот человека оценивается в 20—30 дБ, а крик — в 80 дБ. Болевое ощущение у человека возникает при силе звука 140 дБ. При силе звука 160 дБ начинает разрушаться барабанная перепонка. Человек различает звуковые волны с частотой от 20 Гц до 20 кГц.
Строение уха.Ухо человека делят на наружное, среднее и внутреннее. Звукопроводящей средой для уха является воздух, колебания которого передаются через наружный слуховой проход к барабанной перепонке. Барабанная перепонка, разделяющая наружное и среднее ухо, через цепочку трех взаимосвязанных косточек — моло
точка, наковальни и стремечка — передает колебания внутреннему уху. Внутреннее ухо находится в височной кости и благодаря своей форме называется улиткой. В улитке расположен кортиев (спиральный) орган, а в нем — собственно рецепторные, волосковые клетки (фонорецепторы) (рис. 6.7).
Рис. 6.7.Поперечный разрез через внутреннее ухо (вверху] и строение кортиева органа (внизу)'.
1 — нервные волокна, идущие к кортиеву каналу; 2— внутренние волосковые клетки: 3— покровная мембрана; 4— слой миелина; 5— капсула ганглия; 6— спиральный ганглий; 7— спиральная артерия; 8— limbus spiralis; 9— рейснерова мембрана; 10— вестибулярная лестница; 11 — основная мембрана; 12— кортиев туннель; 13— секреторный эпителий; 14— кортиев орган; 15— спиральная связка; 16— барабанная лестница; 17— наружные волосковые клетки
Механизмы слухового восприятия.Рукоятка молоточка вплетена в барабанную перепонку. Другой его конец сочленен с наковальней, которая сочленена со стремечком. Стремечко прилегает к мембране овального окна. Поскольку площадь барабанной перепонки больше площади овального окна, это приводит к усилению звуковых волн. Сила звука регулируется сокращением мышцы, натягивающей барабанную перепонку и фиксирующей стремечко. В барабанной полости благодаря сообщению евстахиевой трубы с полостью глотки поддерживается давление, равное атмосферному. При глотании слуховая труба открывается и в результате давление в барабанной полости становится равным атмосферному.
Внутреннее ухо — улитка — спирально закрученный костный канал, имеющий 2,5 завитка, который разделен основной мембраной и мембраной Рейснера на три узкие лестницы (рис. 6.7). Верхний канал (вестибулярная лестница) начинается от овального окна и соединяется с нижним каналом (барабанная лестница) через отверстие в верхушке — геликотрему. Оба канала заполнены перилимфой. Между этими каналами расположен средний канал (средняя лестница), заполненный эндолимфой. Внутри среднего канала на основной мембране располагается орган Корти с волосковыми рецепторными клетками, составляющими периферический отдел слухового анализатора. Основная мембрана вблизи овального окна уже, чем на верхушке улитки. Над кортиевым органом лежит покровная (текториальная) мембрана, которая соприкасается с волосковыми клетками, при этом энергия звуковых волн без потери информационного смысла трансформируется в нервные импульсы.
Фонорецепторы кортиева органа являются вторичночувствую- щими рецепторами. Афферентные биполярные слуховые нейроны находятся в спиральном ганглии. От каждой клетки спирального ганглия один отросток идет на периферию к волосковым клеткам кортиева органа, а другой — в составе слухового нерва направляется в ЦНС.
Звук вызывает колебания эндолимфы улиткового протока (средней лестницы) попеременно в сторону вестибулярной и в сторону барабанной лестниц. Результатом такого движения является смешение основной и покровной мембран кортиева органа относительно друг друга. Так как цилии волосковых клеток плотно соприкасаются с покровной мембраной, сгибание цилий вызывает возбуждение волосковых клеток. При этом в волосковых клетках возникает рецепторный потенциал, который вызывает высвобождение медиатора. Медиатор действует возбуждающим образом на постсинаптическую ме
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|