- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
16! 3.3. Принцип лимитирующих факторов
14! 4. 1. Принцип экологического оптимума
На градиенте любого экологического фактора распростране-
ние вида ограничено пределами толерантности, за которыми ор-
ганизм существовать не может (рис. 4).
Между этими пределами
есть отрезок, на котором условия для конкретного вида наиболее
благоприятны и потому формируются самая большая биомасса и
высокая плотность популяции. Это его экологический оптимум
Слева и справа от оптимума условия для жизни вида менее бла-
гоприятны. Это зоны пессимума, т. е. угнетения организмов, ко-
гда падает плотность популяции и вид становится наиболее уяз-
вимым к действию неблагоприятных экологических факторов
(включая и влияние человека).
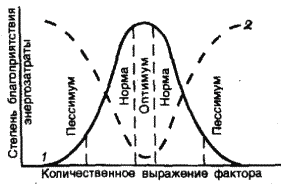
Рис. 4. Схема распределения вида на градиенте среды
(1 – степень благоприятствования условий среды для организма,
2 – величина энергозатрат на поддержание жизнедеятельности)
В зоне оптимума жизнь организма наиболее комфортна, и он
затрачивает на ее поддержание минимальное количество энергии.
В зонах пессимума для поддержания жизнедеятельности приходит-
ся тратить энергии много больше и включать специальные «меха-
низмы выживания». Например, для согревания в холоде тепло-
кровные животные затрачивают энергию, накопленную в жировых
тканях. Растения в условиях пессимума затрачивают значительную
часть продуктов фотосинтеза на дыхание и медленно растут.
Симметричная колоколовидная кривая, показанная на рис. 4,
является достаточно схематизированным приближением к реаль-
ной картине отношений видов к факторам среды. Часто распреде-
ления видов имеют асимметричный характер, т. е. кривая распреде-
ления более пологая с одной стороны и более крутая – с другой.
Кроме того, иногда распределения могут быть двухвершинными
или даже многовершинными, так как на некоторых отрезках гради-
ента вид может быть подавлен более сильными конкурентами.
Разные виды имеют разные амплитуды распределения по
градиентам факторов среды. Виды с узкой амплитудой называ-
ются стенобионтами, с широкой – эврибионтами (рис. 5).
15! 4. 2. Принцип индивидуальности экологии видов
и экологические группы
Каждый вид индивидуально распределяется по экологиче-
ским факторам, кривые распределений разных видов перекрыва-
ются, но их оптимумы различаются. По этой причине при изме-
нении условий среды в пространстве (например, от сухой верши-
ны холма к влажному логу) или во времени (при пересыхании
озера, при усилении выпаса, при зарастании скал) состав экоси-
стем изменяется постепенно. Известный российский эколог
Л. Г. Раменский сформулировал этот закон образно: «Виды – это
не рота солдат, марширующих в ногу».
Индивидуальность экологии видов – это то свойство, которое
«крайне неудобно» для практического использования данных о
связи видов и условий среды. По этой причине для удобства ис-
пользования в практике (т. е. из прагматических соображений)
знание об индивидуальных особенностях видов упрощается (ре-
дуцируется) до системы экологических групп. Такие группы объ-
единяют не тождественные по экологии виды, а виды с близким
отношением к одному или нескольким факторам среды.
Для выделения экологических групп градиент среды разбива-
ется на некоторое число классов, и к одной экологической группе
относятся виды, которые имеют зоны оптимума в этом классе.
16! 3. 3. Принцип лимитирующих факторов
Суть этого принципа, связанного с именем Ю. Либиха, заключается в том, что наиболее важным для распределения вида является тот фактор, значения которого находятся в минимуме или в максимуме.
Как уже отмечалось, наличие в составе комплексного градиента лимитирующего фактора делает этот градиент ведущим, т. е. в большей степени, чем другие, определяющим состав и продуктивность экосистем и состояние популяций формирующих их видов. Так в зоне тундры основу ведущего комплексного градиента составляет количество тепла, так как влаги там достаточно, а обеспеченность элементами минерального питания также зависит от тепла: чем теплее субстраты, тем активнее идет в них процесс минерализации органического вещества и меньше накапливается неразложившихся остатков растений.
В зоне тайги лимитирующим фактором является обеспеченность почв питательными элементами. Почвы, формирующиеся на карбонатных породах, которые богаты кальцием и другими минеральными элементами, позволяют формироваться очень продуктивным сообществам. Однако в условиях тайги, особенно в южной ее части, возрастает роль второго ведущего комплексного градиента, связанного с увлажнением.
В лесостепной и степной зонах ведущий комплексный градиент формируют экологические факторы, связанные с лимитирующим фактором увлажнения, так как почвы в этих зонах (черноземы) богаты питательными элементами. Особенно остро влияние лимитирующего фактора проявляется в годы засух. На этот комплексный градиент может накладываться влияние выпаса (как отмечалось, выпас – это также комплексный градиент), а в южной части степной зоны – комплексного градиента засоления почвы.
В водных экосистемах для большинства входящих в их состав организмов лимитирующими факторами являются содержание кислорода и фосфора, а для растений, кроме того, – свет.
Для многих животных в условиях умеренного климата лимитирующим фактором является глубина снежного покрова. Свободное перемещение по глубокому снегу свойственно сравнительно небольшому числу видов, имеющих «лыжи» (заяц-беляк, заяц-русак, белая куропатка) или отличающихся «длинноногостью» (лось). Глубокий снег является препятствием для перемещения волка и кабана. Этот фактор определяет границы коневодства с круглогодичным содержанием животных на пастбищах. При глубоком снеге (в период зимнего выпаса – тебеневки) лошади не могут использовать корм.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|