- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Нуклеиновые кислоты.
Лекция№-1.
Тема: «Цитологические основы наследственности».
Основные теоретические положения
Время жизни клетки с момента ее образования до деления составляет клеточный цикл. Клеточный цикл делится на стадии: митоз или собственно деление клетки и интерфаза – период между делениями. Самой длительной стадией интерфазы является период относительного покоя клетки – пресинтетическая стадия G1, ее продолжительность может значительно варьировать. После G1 начинается очень важная синтетическая стадия S, в процессе которой происходит репликация ДНК. Далее следует подготовка к митозу – постсинтетическая стадия G2 и сам митоз – стадия М.
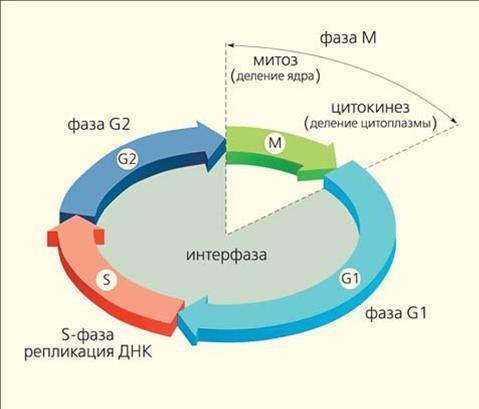
Рисунок 7. Схема клеточного цикла7
Ключевая роль в митозе принадлежит хромосомам – таким структурам в ядрах клеток, которые в период деления отчетливо видны при световой микроскопии и использовании специфических методов окрашивания. Окрашивающееся вещество хромосом называется хроматином.
Набор хромосом одной клетки называется кариотипом. Число и морфология хромосом относятся к видовым признакам.
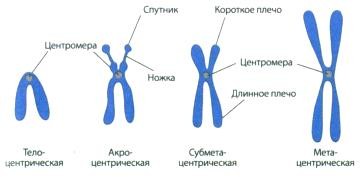
Рисунок 8. Типы хромосом8
В каждой хромосоме есть важный функциональный участок, который называется центромерой. Центромера разделяет хромосому на два плеча, которые называются теломерами. Хромосомы делят на акроцентрики, метацентрики и субметацентрики при локализации центромеры соответственно на конце хромосомы, посредине и в промежуточном положении. Выделяют также телоцентрические хромосомы, короткое плечо у которых отсутствует.
В соматических клетках высших каждая хромосома представлена двумя копиями, то есть диплоидным набором. И только в половых клетках наблюдается одинарный или гаплоидный набор хромосом, образующийся в результате процесса мейоза.
Митоз, в свою очередь, также делится на стадии. На стадии профазы происходит исчезновение ядерной мембраны, конденсация или уплотнение хромосом за счет их спирализации, миграция центриолей к противоположным полюсам, приводящая к поляризации клетки, и формирование веретена деления.
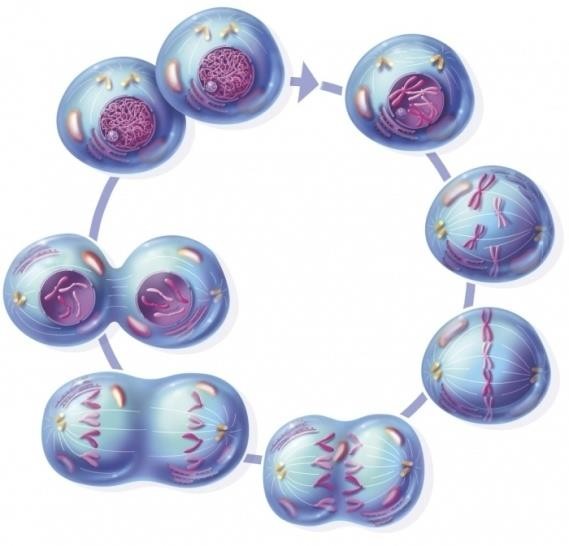 Рисунок 9. Митоз
Рисунок 9. Митоз
В период метафазы центромеры располагаются по экватору клетки перпендикулярно оси веретена деления. На стадии анафазы происходит разделение центромер, хроматиды превращаются в самостоятельные хромосомы и, увлекаемые центромерами, начинают двигаться к противоположным полюсам клетки по нитям веретена деления. На заключительной стадии – телофазе – происходит деспирализация хромосом, исчезает веретено деления, формируется ядерная мембрана и происходит разделение цитоплазмы ( цитокенез ).
Мейоз происходит только при образовании половых клеток, и он включает два клеточных деления: мейоз I или редукционное деление и мейоз II (эквационное деление). Во время профазы мейоза I гомологичные хромосомы коньгируют (сливаются) друг с другом по всей длине, образуя бивалент. В это время может происходить обмен участками между несестринскими хроматидами – кроссинговер или гомологичная рекомбинация. В точке рекомбинации образуется видимая в световой микроскоп крестообразная структура – хиазма. Обмен происходит только между двумя из
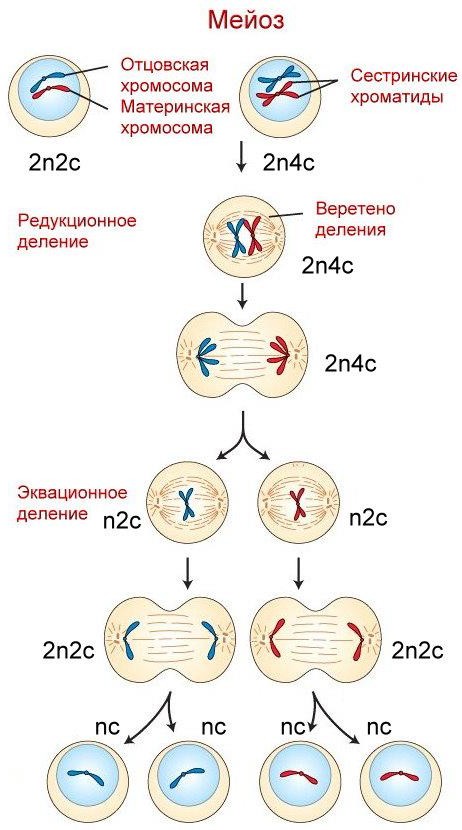
четырех хроматид. На стадии метафазы биваленты выстраиваются в экваториальной плоскости, при этом ценромеры случайно ориентируются относительно полюсов клетки. На стадии анафазы гомологичные хромосомы отделяются друг от друга и начинают двигаться к противоположным полюсам. При этом расщепления центромеры не происходит, и сестринские хроматиды оказываются связанными. Однако они могут быть уже не идентичны друг другу из-за произошедшего кроссинговера. Таким образом, в процессе мейоза I из одной диплоидной клетки образуются две гаплоидные. Промежуток между первым и вторым делениями мейоза называется интеркинезом. Важно подчеркнуть, что на этой стадии не происходит удвоения хроматид.
Рисунок 10. Мейоз
В профазе мейоза II восстанавливается веретено деления, хромосомы располагаются в экваториальной плоскости. В анафазе II происходит расщепление центромер, и хромосомы двигаются к противоположным
полюсам. Таким образом, на один акт удвоения хромосом приходятся два последовательных цикла деления клетки. После завершения телофазы II диплоидная родительская клетка делится на четыре гаплоидные половые клетки, причем образовавшиеся гаметы не идентичны друг другу – фрагменты материнских и отцовских хромосом находятся в них в различных комбинациях.
Нуклеиновые кислоты.
В начале 50-х годов ХХ века было доказано, что единицей наследственности и изменчивости является ген, материальной основой которого является дезоксирибонуклеиновая кислота (ДНК).
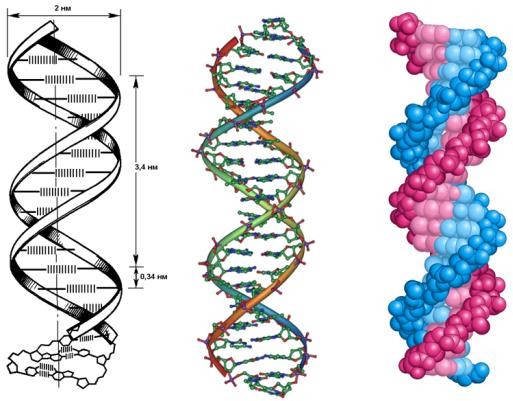
Структура молекулы ДНК была расшифрована Дж. Уотсоном, Ф. Криком и М. Уилкинсом в 1953 г. Она представляет собой (рис. 1) две спирально закрученные антипараллельные (напротив конца 3/ одной цепи располагается 5/ конец другой) полинуклеотидные цепи.
Рисунок 1. Структура молекулы ДНК1
Мономерами ДНК являются нуклеотиды (рис. 2), в состав каждого из них входят:
1. пятиуглеродный сахар – дезоксирибоза;
2. остаток фосфорной кислоты;
3. одно из четырех азотистых оснований (пуриновые: аденин и гуанин, пиримидиновые: тимин и цитозин ).
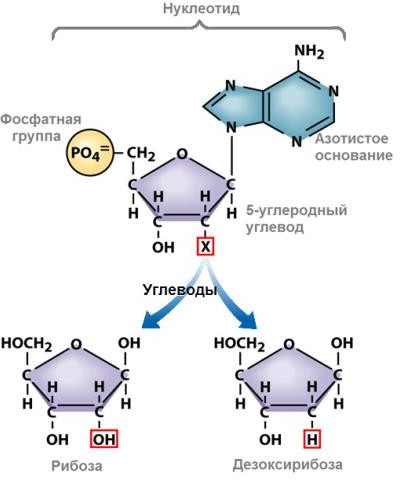
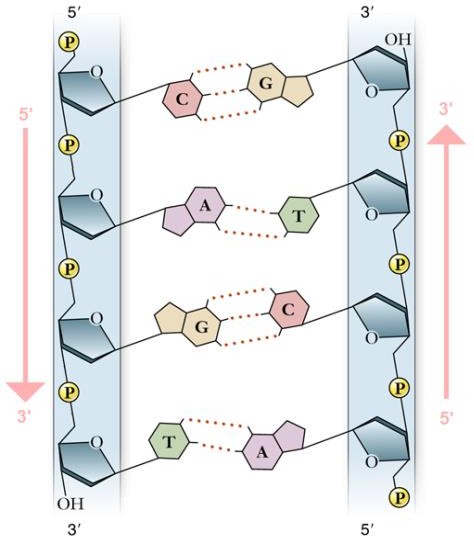
Рисунок 2. Строение нуклеотида2 Рисунок 3. Вторичная структура
молекулы ДНК3
Нуклеотиды соединяются в цепочку путем образования ковалентных (фосфодиэфирных) связей между дезоксирибозой одного и остатком фосфорной кислоты другого нуклеотида. Азотистые основания присоединяются к дезоксирибозе и образуют боковые радикалы. Между азотистыми основаниями цепочек ДНК устанавливаются водородные связи: две
– между аденином и тимином, три – между гуанином и цитозином (рис. 3). Строгое соответствие (взаимодополнение) нуклеотидов друг другу в парных цепочках ДНК (А – Т, Г – Ц) называется комплементарностью.
ДНК является хранителем генетической информации во всех клетках про- и эукариот.
Рибонуклеиновая кислота (РНК) в отличие от ДНК содержит сахар – рибозу. Также в молекуле ДНК не встречается азотистое основание – тимин, в
состав молекулы входит другое пиримидиновое основание – урацил. Необходимо также отметить, что молекула РНК в отличие от ДНК представлена одинарной цепочкой.
В клетках существуют три типа РНК: информационная, транспортная и рибосомальная.
Информационная РНК (и-РНК) представляет собой копию определенного участка ДНК и выполняет функции переносчика генетической информации от ДНК к месту синтеза белка (рибосомы) и непосредственно участвует в сборке его молекул.
Транспортные РНК (т-РНК) транспортируют аминокислоты из цитоплазмы в рибосомы.
Рибосомальная РНК (р-РНК) входит в состав рибосом. Считают, что р- РНК обеспечивает определенное пространственное взаиморасположение и-РНК и т-РНК.
Репликация молекул ДНК происходит в синтетический период интерфазы при участии комплекса ферментов (рис. 4).
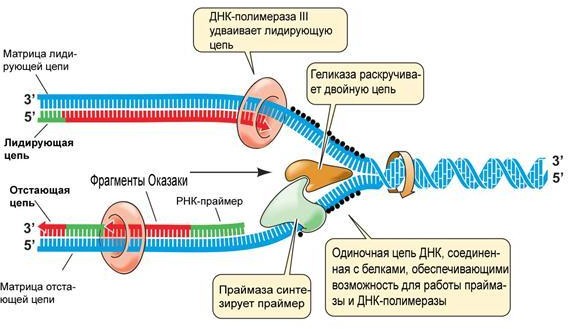
Рисунок 4. Схема репликации молекулы ДНК4
Каждая из двух цепей «материнской» молекулы служит матрицей для
«дочерней». Строит дочернюю молекулу фермент ДНК-полимеразы согласно принципу комплементарности. После репликации вновь синтезированная молекула ДНК содержит одну «материнскую» цепочку, а вторую –
«дочернюю», вновь синтезированную (полуконсервативный способ).
Транскрипция – процесс синтеза молекулы и-РНК, происходящий в ядре. Фермент РНК-полимераза подходит к молекуле ДНК и разрывает водородные связи, после чего молекула ДНК раскручивается на 2 цепочки. Одна из цепей ДНК является кодирующей. Она начинается с 3/ конца, так как фермент РНК- полимераза движется именно в этом направлении, и транскрипция осуществляется в направлении 3/ → 5/, а иРНК образуется в направлении 5/ → 3/. Из свободных нуклеотидов РНК фермент строит молекулу и-РНК по принципу комплементарности азотистых оснований нуклеотидов (аденину ДНК соответствует урацил РНК, тимину ДНК – аденин РНК, гуанину ДНК – цитозин РНК, цитозину ДНК – гуанин РНК). Генетическая информация молекул ДНК преобразовалась в последовательность нуклеотидов молекулы и- РНК, которая затем выходит из ядра и направляется к рибосомам для процесса синтеза белка.
Трансляция – перевод генетической информации с языка последовательности нуклеотидов на язык последовательности аминокислот, (расшифровка генетического кода). Происходит в цитоплазме на рибосомах. Участвуют: и-РНК, аминоацил-т-РНК, р-РНК, ферменты. Считывание информации с и-РНК осуществляется в направлении 5/ → 3/. Трансляция состоит из трех этапов: инициация, элонгация и терминация (рис 5. ).
Инициация – объединение малой субъединицы рибосом, инициирующего триплета и-РНК (АУГ), метионин-аминоацил-т-РНК и большой субъединицы рибосомы.
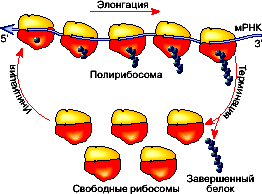
Рисунок 5. Схема трансляции белка5
Элонгация – наращивание полипептида. Эта фаза включает все реакции с момента образования первой пептидной связи до присоединения последней аминокислоты.
На заключительном этапе трансляции ( терминация ) рибосома доходит до одного из «nonsens» кодонов иРНК и синтез полипептида прекращается.
Информация о первичной структуре белковой молекулы закодирована последовательностью нуклеотидов ( генетический код ) в соответствующем участке молекулы ДНК – гене.
Свойства генетического кода:
1. Однозначность, т. е. каждый триплет (три нуклеотида) кодирует только одну аминокислоту.
2. Избыточность – некоторые из аминокислот будут кодироваться 2, 3, 4 или 6 триплетами (валин кодируется 4 триплетами, а серин – 6).
3. Неперекрываемость – одновременно 1 нуклеотид входит в состав только одного триплета.
4. Универсальность – у всех организмов одинаковые триплеты кодируют одинаковые аминокислоты.
5. Однонаправленность – код читается только в одном направлении.
6. Наличие нонсенс (стоп)-кодонов – триплетов, которые не кодируют аминокислоты. Когда рибосома в процессе трансляции доходит до таких кодонов, то синтез белка прекращается. В молекуле РНК – УАА, УГА, УАГ.
7. Непрерывность (без знаков препинания) – при выпадении одного нуклеотида в процессе считывания его место занимает нуклеотид из соседнего кодона. Стартовыми кодонами в молекуле и-РНК являются АУГ и ГУГ.
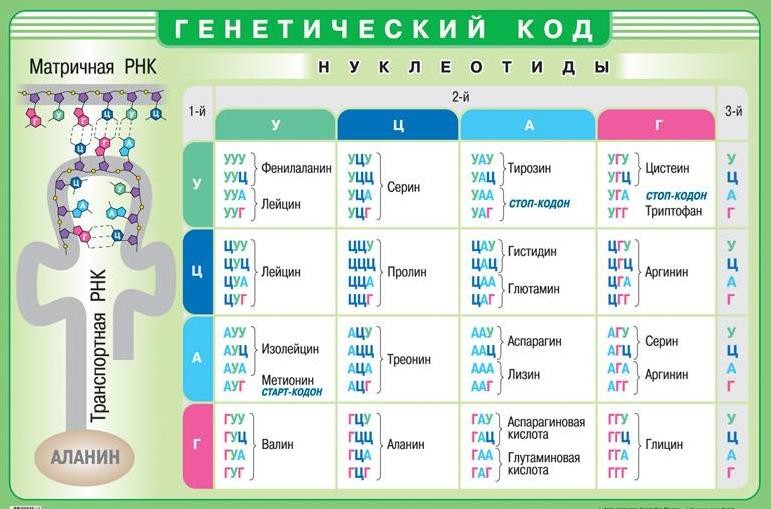
Рисунок 6. Таблица генетического кода
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|