- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Анализ подвидовой принадлежности выборки из Бурзянского района РБ
Анализ подвидовой принадлежности выборки из Бурзянского района РБ
В таблице 1 представлены данные по полиморфизму микросателлитных локусов и локуса COI-COII мтДНК в выборке из Бурзянского района. Аллельный вариант Q был зафиксирован только в одной из 10 семей. В семье № 220 был выявлен аллельный вариант PQQQ.
Таблица 1. Аллели COI-COII и микросателлитных локусов ap243, 4a110, A24, A8, A43, A113, A88, Ap049, A28 в исследуемой выборке рабочих пчел (Республика Башкортостан, Бурзянский район, д. Новоусманово, Сайфутдинова Фируза Талгатовна, от 08. 10. 2021 г. )
|
№ семьи |
COI-II |
Ap243 |
4a110 |
А24 |
A8 |
A43 |
A113 |
A88 |
Ap049 |
A28 |
| PQQ | ||||||||||
| PQQ | ||||||||||
| Q | ||||||||||
| PQQ | ||||||||||
| PQQQ | ||||||||||
| PQQ | ||||||||||
| PQQ | ||||||||||
| PQQ | ||||||||||
| PQQ | ||||||||||
| PQQ |
На основе данных по полиморфизму микросателлитных локусов был рассчитан уровень интрогрессии генофонда ветви С (таблица 2, жирным шрифтом выделены семьи, принадлежащие подвиду A. m. mellifera). Для выборки из Бурзянского района среднее значение уровня интрогрессии составило 5. 4%. Не смотря на наличие аллельного варианта Q в семье №53, она имеет низкий уровень гибридизации. Возможной причиной этому является благоприятный трутневый фон.
Таблица 2. Уровень интрогрессии генов подвидов эволюционных ветвей М и С в семьях пчел, рассчитанный на основе анализа полиморфизма микросателлитных локусов ap243, 4a110, A24, A8, A43, A113, A88, Ap049, A28
|
№ семьи |
COI-II |
Доля С |
Доля М |
|
PQQ |
0. 02 |
0. 98 | |
|
PQQ |
0. 02 |
0. 98 | |
| Q | 0. 13 | 0. 87 | |
|
PQQ |
0. 06 |
0. 94 | |
|
PQQQ |
0. 03 |
0. 97 | |
|
PQQ |
0. 02 |
0. 98 | |
|
PQQ |
0. 02 |
0. 98 | |
|
PQQ |
0. 03 |
0. 97 | |
|
PQQ |
0. 10 |
0. 90 | |
|
PQQ |
0. 11 |
0. 89 | |
|
Среднее |
0. 054 |
0. 946 |
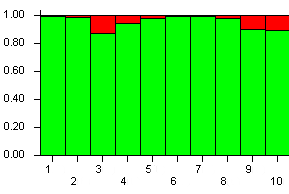
Рисунок 1 Генетическая структура выборки из Бурзянского района, зеленым цветом обозначена доля эволюционной ветви М, красным – ветви С.
Таким образом, исследуемые семьи из Бурзянского района принадлежат к подвиду Apis mellifera mellifera (доля М > 0. 88), за исключением семьи № 53. В семье № 53 хороший отцовский генофонд, тем не менее, в ней необходимо провести замену матки.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|