- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Гистоновый код: искусство упаковки с хвостами
Метилирование ДНК. Слева — основания цитозин и гуанин всегда вместе образуют ступеньку в молекуле ДНК, так как атомы водорода (Н) каждого из оснований притягивают их друг к другу (водородные мостики). Фермент ДНК-метилтрансфераза (DNMT) связывается с цитозином, когда хочет прикрепить метильную группу (СНЗ). Справа — модель метилированного участка ДНК. Метилирование приводит к тому, что наследственную информацию на этом участке прочитать невозможно.
Темой нескольких берлинских докладов были вещества под названием ДНК-метилтрансферазы (сокращенно DNMT). Известно 4 типа ДНК-метилтрансфераз. Название позволительно забыть, но функция этих веществ настолько важна, что ее стоит запомнить: ДНК-метилтрансферазы встраивают в наследственное вещество крохотные, но чрезвычайно действенные задвижки и таким образом выключают гены. Следовательно, они имеют право решать, какие белки клетка синтезирует, а какие нет.
Задвижки — это метильные группы, то есть простейшие химические структуры, состоящие из одного атома углерода и трех атомов водорода. При необходимости ДНК-метилтрансферазы прочно прикрепляют эти группы к одной из букв генного текста, точнее — к основанию цитозин. Если этот цитозин, как выражаются химики, «метилирован», то считывающие генный текст белки не смогут присоединиться к двойной спирали ДНК. Соответствующий ген выключен.
В клетках млекопитающих такими задвижками снабжены больше двух третей соответствующих участков. Речь идет лишь о тех местах, где наблюдается большое количество пар цитозиновых оснований, расположенных на обеих нитях ДНК с некоторым смещением относительно друг друга. Это происходит, когда основания цитозин и гуанин, вместе образующие ступеньку ДНК, часто чередуются (CpG-острова). Тогда генный текст одной цепочки выглядит так: CGCGCG, а другой — так: GCGCGC. В этом случае ДНК метилирована по обеим цепочкам. Благодаря этому хитрому механизму клетка при делении передаст по наследству обеим своим дочкам модель метилирования собственной ДНК. То есть каждая дочка наследует, как описано выше, одну из двух материнских цепочек ДНК. Если бы метилирована была лишь одна цепочка, какая-то из дочерних клеток не получила бы эту информацию, а выключенные гены вновь были бы активированы.
Модель метилирования соответствует типу клетки и четко определяет, что клетка может, а что — нет. В оплодотворенной яйцеклетке метилирование практически отсутствует, поскольку из нее развиваются все типы соматических клеток. Только когда миллиарды дочерних клеток разделяются по назначению, ДНК-метилтрансферазы встраивают одну задвижку за другой.
Как именно это происходит, пока не изучено. Однако бременский биохимик и специалист по ДНК-метилтрансферазам Альберт Йельч предполагает, что клетка отключает те участки ДНК, которые ей в данный момент не нужны. «Идея заключается в том, что модели метилирования стабилизируют и фиксируют активность генома, — говорит ученый. — Если тот или иной ген был активирован, он и дальше будет настроен на активность».
Например, этот простой принцип объясняет, почему эмбриональная клетка в будущей кожной ткани сама программируется как клетка кожи и передает эту программу по наследству своим дочкам, так что впоследствии из нее могут получаться только клетки кожи. Естественно, любая еще не дифференцированная клетка по этой модели может развиваться в любой вид ткани. Решающее значение имеют при этом сигналы организма, активизирующие определенную программу внутри клетки. В заключение метильные группы обеспечивают клетке долгосрочное запоминание этой программы.
Так вкратце можно описать то, что эпигенетики понимают под «клеточной памятью». Эпигенетическая программа замораживает модели активации генов и таким образом запоминает информацию. Данный принцип также позволяет объяснить, почему на геном влияют не только программы биологического развития, но и повседневные внешние воздействия. Последние посредством биохимических сигналов организма тоже могут побудить клетки считывать или подавлять гены, тем самым предоставляя метилтрансферазам возможность встраивать в ДНК новые задвижки.
Гистоновый код: искусство упаковки с хвостами
Разумеется, на берлинской конференции по эпигенетике я узнал много нового. Даже из докладов. Например, мне стало известно, что у многих белков есть хвост. У некоторых даже два. Это, разумеется, не отросток позвоночника, как у мышей или кошек, а конец или начало цепочки, из которой состоит белок. Хвост торчит из конструкции, получившейся в результате синтеза молекулы.
Конечно, можно посчитать, что природа пошутила, снабдив некоторые из своих кирпичиков хвостами. Однако в природе практически все имеет смысл, вот и хвосты белков не исключение — они играют решающую роль в программировании наследственного материала. Чтобы объяснить это нагляднее, мне придется уменьшиться в миллион раз и совершить путешествие в невообразимо миниатюрный наномир соматических клеток.
Внутри клетки кипит жизнь: там есть белковые фабрики, электростанции, клеточный скелет, множество снующих туда-сюда молекул-медиаторов и молекул-транспортеров, а также многое другое. Но в центре действия — неподвижное клеточное ядро, толстое и круглое, словно пчелиная матка в улье. Размер ядра — одна сотая миллиметра, и это самая крупная часть клетки. Через одну из многочисленных пор я попадаю внутрь ядра. Обычно этим путем наружу проникает транспортная РНК. Она доставляет на белковые фабрики считанные с ДНК «монтажные схемы».
У меня перехватывает дыхание: внутри ядра тоже повсюду снуют белки, оседающие в разных местах тонюсенькой ниточки. Белки участвуют в считывании генетического кода, а ниточка — не что иное, как одна из 46 молекул ДНК — хромосом, содержащих этот код. Толщина хромосомы — всего 0, 3 нанометра, то есть треть одной миллионной миллиметра, зато длина ДНК в совокупности составляет легендарные два метра.
Если бы эти нити по всей своей длине беспорядочно переплетались, воцарился бы хаос. (Попробуйте-ка уложить два метра ниток в коробочку размером с клеточное ядро. ) К счастью, существуют структуры, отвечающие за порядок: я вижу округлые образования из нескольких белков, на которые ДНК наматывается, словно кабель на барабан. Одновременно множество подобных барабанов связываются в цепочку, напоминающую жемчужное ожерелье.
Издалека это ожерелье выглядит очень изящно. И неудивительно, ведь поперечник нити — всего одиннадцать миллионных миллиметра. Но в некоторых местах заметны утолщения. Я подхожу поближе и обнаруживаю, что именно там барабаны вместе с нитью ДНК делают еще один виток. На этот раз они образуют спираль. В результате нить еще раз заметно укорачивается и утолщается, образуя 30-нанометровую фибриллу.
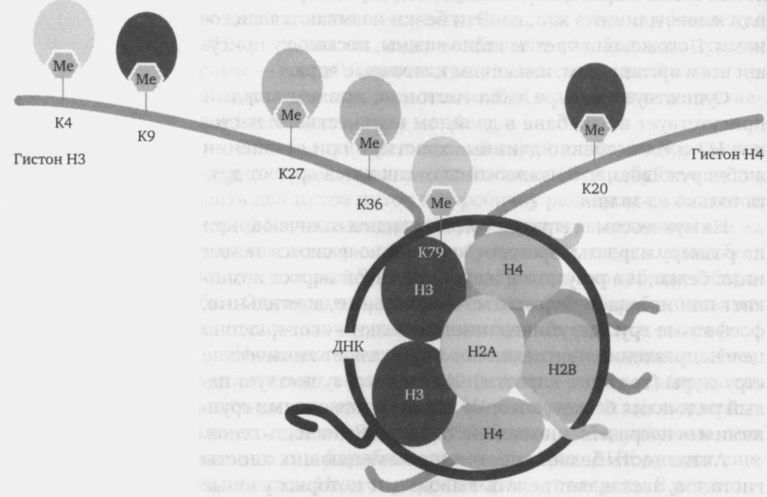
Это спирально закрученное переплетение белков и ДНК теперь в сто раз толще изначального наследственного вещества. Биологи называют такое переплетение хроматином — независимо от того, насколько плотно оно упаковано. В сильно скрученном состоянии оно называется гетерохроматином. Когда клетка находится в процессе деления, например, если ткань, в которую она входит, должна расти, гетерохроматин скручивается особенно плотно, чтобы отдельные хромосомы могли распутаться и распределиться по дочерним клеткам. Вернемся к многочисленным барабанам, на которые нить ДНК во многих местах наматывается в два-три оборота. Эти так называемые нуклеосомы на первый взгляд одинаковы. Они состоят из восьми шаровидных белков, из которых торчат более или менее длинные хвосты. Эти белки называются гистонами. Похоже, они чрезвычайно важны, поскольку присущи всем организмам, имеющим клеточные ядра.
Существует четыре типа гистонов, причем каждый присутствует в барабане в двойном количестве. У гистонов Н3 и Н4 особенно длинные хвосты, и при сравнении я обнаруживаю, что нуклеосомы отличаются друг от друга только из-за них.
На нуклеосомах в разных местах видны отличающиеся по размеру наросты. Время от времени появляются те или иные белки, и в результате их работы такой нарост возникает или исчезает. Ферменты — метильные, ацетильные, фосфатные группы, убиквитиновые белки — совершенно целенаправленно присоединяют или удаляют химические структуры (те самые наросты). Кроме того, существует целый ряд малых белков, которые связываются с этими группами и непосредственно воздействуют на активность генов.
Активность белков, постоянно изменяющих хвосты гистонов, заставляет делать выводы, от которых ученые отказывались всего 15 лет назад. Теперь это общепризнанное мнение: благодаря изменчивости гистоновых хвостов эпигенетический код может оказывать удивительно детальное, тонко дифференцированное и многоаспектное влияние на назначение и свойства клетки.
Заклинание звучит так: модификация гистонов. В зависимости от того, какой нарост и какой малый белок присоединяются к тому или иному участку хвоста, пространственная структура гистона строго определенным образом меняется. В результате нить ДНК либо плотнее, либо свободнее прилегает к барабану, так что ряду дополнительных белков, выполняющих важные функции, легче или труднее к ней присоединиться.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|