- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Опыт Гриффитса.. Опыт Херши и Чейза.
Опыт Гриффитса.
Работал с двумя штаммами пневмококка, которые отличались по внешнему виду и болезнетворным свойствам. Штамм S – от него мыши сразу погибали. Штамм R – при введении гибели мышей не наступало. Он заметил, что при смеси убитых вирулетных и невирулетных мышы погибали. Это доказало процесс транформации.
Трансформация – включение чужеродной ДНК в бактериальную клетку. Это перенос наследственной информации от одной клетки прокариотов к другой посредством ДНК бактерии – донора или клетки – донора. (Гриффитс, 1928).
Опыт Херши и Чейза.
Они выяснили, что не все вирусы убивают бактериальную клетку. Существуют умеренные фаги. Они могут быть и вирулентными и объединятся с геномом бактериальной клетки. При этом попадая в другие бактерии вирусы привносят чужие генетические данные. Так доказан процесс трансдукции.
Трансдукция заключается в том, что вирусы, покидая бактериальные клетки, в которых они паразитировали, могут захватывать с собой часть их ДНК и, перемещаясь в новые клетки, передавать новым хозяевам свойства прежних.
Доказательства генетической роли ДНК были получены в ряде опытов по заражению бактериальных клеток вирусами. Бактериофаг состоит из белковой капсулы правильной геометрической формы и молекулы нуклеиновой кислоты, свернутой в виде спирали. Фаг прикрепляется своими отростками к клеточной оболочке, с помощью ферментов разрушает участок клеточной мембраны и чрез образовавшееся отверстие вводит свою ДНК в клетку и т. д.
Когда белок фага был помечен радиоактивной серой 35S, а ДНК – радиоактивным фосфором 32Р оказалось, что вновь образованные фаги содержали только радиоактивный фосфор, которым была помечена ДНК. Эти опыты наглядно показали, что генетическая информация от внедрившегося фага его потомкам передается только проникающей в клетку нуклеиновой кислотой, а не белком, содержащимся в капсуле вируса.
Опыт Френкель-Конрата.
Им удалось доказать, что чистая НК вируса табачной мозаики может заражать растения. И удалось создать гибрид из вирусов, в котором белковый футляр принадлежал одному, а НК другому вирусу. В таких случаях генетическая информация гибридов в точности повторяла генетическую информацию вируса, чья НК находилась в гибриде.
Современные представления о генетическом коде. Опыт Ниринберга. Синтез специфических белков. Репликация, транскрипция, трансляция. Транскрипция 4х-значного кода первичной генетической информации в 20-значный аминокислотный код белков. Виды и структура т-РНК, р-РНК, и-РНК. Мультимерная организация белков (гемоглобин человека: HbA, HbA2, HbS, HbF).
Полуконсервативная репликация ДНК.
Полуконсервативный механизм репликации предполагает разделение цепей исходной молекулы ДНК. При этом каждая материнская цепь является матрицей для синтеза дочерней цепи. Таким образом, при репликации образуются две молекулы ДНК, каждая из которых включает одну материнскую цепь и одну новую — дочернюю. 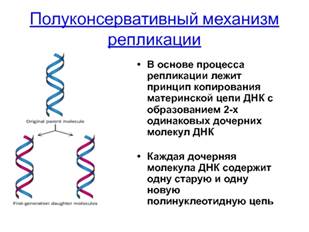
ДНК-полимеразы.
- это Фермент, участвующий в репликации ДНК. Ферменты этого класса катализируют полимеризацию дезоксирибонуклеотидов вдоль цепочки нуклеотидов ДНК, которую фермент «читает» и использует в качестве шаблона. Тип нового нуклеотида определяется по принципу комплементарности с шаблоном, с которого ведётся считывание. Собираемая молекула комплементарна шаблонной моноспирали и идентична второму компоненту двойной спирали.
Вилка репликации ДНК.
Процесс репликации происходит в специальных структурах, названных вилками репликации. Схематическое устройство репликативной вилки E. coli представлено на рис. 1. То, что две нити молекулы ДНК расположены антипараллельно друг другу, создает ряд проблем для их олдновременной разнонаправленной репликации.
По мере движения вилки одновременно должны синтезироваться две дочерние цепи. Вилка движется в направлении от 5' к 3’ на одной цепи и от 3’ к 5' – на другой. Однако нуклеиновые кислоты синтезируются только от 5'– к 3'-концу. Проблема решается таким образом, что на одной из родительских нитей новая нить синтезируется непрерывно в направлении 5'-3', что совпадает с движением вилки репликации. Это называется лидирующей или ведущей. Другая нить называется отстающей или запаздывающей, так как синтез на ней идет с некоторой задержкой по сравнению с лидирующей нитью. Это связано с тем, что ДНК на этой нити синтезируется также от 5' к 3', но в направлении, противоположном движению вилки, и короткими фрагментами. Благодаря этому разнонаправленный синтез ДНК может осуществляться в рамках одной структуры – репликативной вилки.
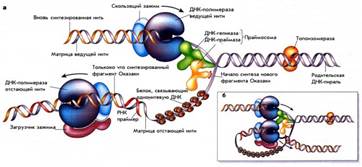
Рис. 1. Схема репликативной вилки.
Длина таких коротких фрагментов у прокариот составляет 1000–2000 пн. По имени открывшего их ученого они были названы «фрагментами Оказаки». По мере движения репликативной вилки концы соседних фрагментов Оказаки соединяются с образованием непрерывной отстающей нити. Для того, чтобы процесс на обеих нитях шел синхронно, полимеразные комплексы лидирующей и отстающей нити связаны между собой, образуя сложную трехмерную структуру (рис. 1, б)
Вилка репликации может двигаться как в одну сторону от точки начала репликации, так и в обе стороны. В зависимости от этого процесс называется однонаправленной или двунаправленной репликацией. Как это выглядит схематически, показано на рис. 2. У эукариот репликация обычно двунапраленная. Также и у E. coli.
Механизмы инициации репликации в точке начала репликации и при образовании фрагментов Оказаки в отстающей цепи в принципе аналогичны, хотя имеются некоторые тонкие различия. В обоих случаях происходит образование коротких РНК-затравок ( праймеров ), комплементарных матричной ДНК, в виде продолжения которых синтезируется новая цепь ДНК. В дальнейшем короткие вставки РНК замещаются сегментами ДНК, отдельные фрагменты Оказаки затем объединяются с образованием непрерывной отстающей нити.
Все живые организмы на Земле обычно делят на прокариот и эукариот (от греч. карион – ядро). Главной особенностью прокариот является отсутствие у них в отличие от эукариот полноценного клеточного ядра, покрытого оболочкой. Генетический материал прокариот расположен в нуклеоиде – примитивном эквиваленте ядра эукариот. Клетки прокариот имеют очень небольшие размеры – около 1 мкм. Объем эукариотических клеток в 800-1000 раз больше объема клеток прокариот. К прокариотам относятся бактерии и археи (или архебактерии), предки которых возникли около 4 млрд лет назад. Эукариоты могут быть как одноклеточными, так и многоклеточными. Они появились на Земле примерно через 500 млн лет после прокариот.
По современным представлениям ДНК-метаболизм у прокариот имеет некоторые отличия от такового у эукариот. Описывая процессы репликации и рекомбинации, мы будем каждый раз подчеркивать эти отличия.
Регуляция репликации ДНК у бактерий.
Бактериальный геном представляет собой кольцо, размером примерно 5 млн. пар нуклеотидов. В этом кольце есть конкретное место, с которого начинается удвоение. Репликация происходит по следующему принципу. С точки начала репликации в две стороны начинают двигаться два комплекса ферментов. Каждый комплекс ферментов способен удваивать сразу обе цепочки ДНК. Область, в которой он находится, называется вилкой репликации. Два комплекса пройдут по кольцу по часовой и против часовой стрелки. Когда они встретятся, репликация будет завершена.
Ключевым ферментом, осуществляющим репликацию, является ДНК-полимераза. Она удваивает ДНК. Как правило, в каждой клетке существует несколько разных ДНК полимераз, для разных целей. В частности, геном бактерий реплицирует ДНК-полимераза III. ДНК-полимераза I и II — это вспомогательные ферменты, участвующие в процессах исправления ошибок в ДНК (репарации). ДНК-полимераз в репликативной вилке должно быть две, поскольку цепочек ДНК тоже две.
Необходимо помнить, что полимераза может синтезировать ДНК только в одном направлении от 5’- к 3’- концу. Эти штрихи берутся от нумерации атомов в сахаре. Дезоксирибоза — пятиатомный сахар. Она является необходимым звеном, образующим сахарофосфатный остов. В образовании этого остова играют роль 3 и 5 атомы углерода. Соответственно, у сахара, у сахарофосфатного остова, у ДНК и РНК есть 3’-конец и 5’- конец. Синтезироваться молекула ДНК может только путем присоединения новых нуклеотидов к растущему 3’-концу. Энергия для синтеза ДНК берется от нуклеотид-3-фосфатов, которые одновременно являются и структурными элементами ДНК. При присоединении нуклеотида отщепляется пирофосфат, который тут же разрушается специальным ферментом (пирофосфатазой) еще на две отдельных молекулы ортофосфорной кислоты, тем самым, делая эту реакцию необратимой.
Почему ДНК-полимераза присоединяет нуклеотиды к 3’-концу молекулы? Может быть, это связано с тем, что при таком синтезе ДНК-полимераза может репарировать собственные ошибки. Если фермент присоединил неправильный нуклеотид, а потом «осознал», что ошибся, то он может просто сделать шаг назад, удалить нуклеотид и сделать все, как было. Если бы синтез шел наоборот, так нельзя было бы сделать. На самом деле это точно не известно и причины могут быть другими.
Поскольку нить ДНК растет в одном направлении, и ферментный комплекс движется в одном направлении, как осуществляется синтез комплементарной цепи?
Дело в том, что вторая цепь ДНК образует петлю и разворачивается так, чтобы обе ДНК-полимеразы в составе ферментного комплекса могли двигаться вместе (рис. 3). В результате, на второй цепи ДНК синтез идет кусочками, с перерывами. Эта цепь называется отстающей, другая — лидирующей. Время от времени фермент перескакивает на какое-то расстояние по цепи ДНК дальше и синтезирует дочернюю цепь до следующего перескакивания. Синтез с отстающей цепи происходит фрагментами. Эти фрагменты названы в честь японского ученого, который их описал, — фрагментами Оказаки. Фрагменты Оказаки сшиваются лигазой.
Понятие о репликоне и репликаторе.
Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации.
Репликация (от лат. replicatio – повторение) – это самовоспроизведение молекулы ДНК, обеспечивающее точное копирование генетической информации и передачу ее от поколения к поколению. Синтез дочерней молекулы ДНК, идет во время синтетической (S) фазы жизненного цикла клетки на матрице родительской молекулы ДНК.
Хромосомная теория.
Хромосомная теория наследственности — теория, согласно которой передача наследственной информации в ряду поколений связана с передачей хромосом, в которых в определённой и линейной последовательности расположены гены. Эта теория сформулирована в начале XX века, основной вклад в её создание внесли американский цитолог. У. Саттон (англ. ) русск., немецкий эмбриолог. Т. Бовери и американский генетик. Т. Морган со своими сотрудниками.
Положения:
1. Хромосомы являются носителями наследственной информации.
2. Каждая хромосома содержит множество генов.
3. Гены, расположенные в одной хромосоме, наследуются совместно (сцепленно) и образуют группу сцепления.
4. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гетерогаметного пола больше на одну).
5. Сцепление аллелей, расположенных в одной хромосоме, может нарушаться за счет кроссинговера.
6. Гены расположены в хромосомах в линейной последовательности.
7. Частота кроссинговера (нарушения сцепления) пропорциональна расстоянию между генами.
Практическая часть
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|