- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Лабораторное работа № 1.. Тема: Строение растительной клетки.. Теоретический материал.
Лабораторное работа № 1.
Тема: Строение растительной клетки.
Цель: познакомиться с многообразием клеток, с клеточным строением живых организмов, значение клетки, изучить строение клеток растений.
Задание: рассмотрите и опишите, что собой представляет клетка, какие функции выполняет органоиды клетки, ее цитоплазма; изучите, что такое тургор клетки.
Теоретический материал.
Клетка представляет собой основную структурную и функциональную единицу живого организма. У одноклеточных она осуществляет все функции, свойственные целому организму: питания, дыхания, накопления и выделения продуктов жизнедеятельности, размножения и т. п. У многоклеточных каждую из этих функций выполняют соответствующие комплексы клеток, т. е. ткани и органы.
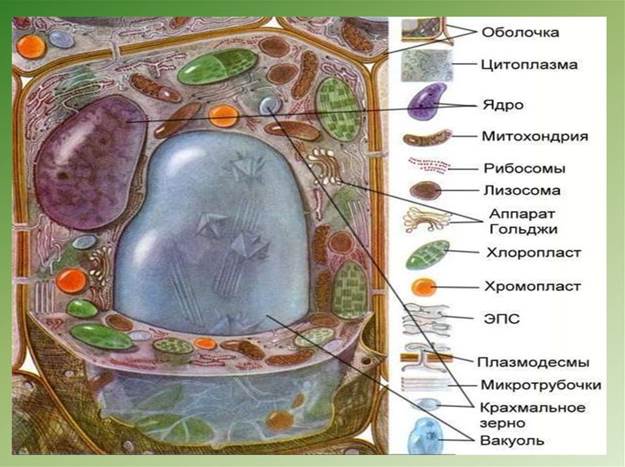
Рис. 1. Строение растительной клетки
По форме клетки могут быть овальные, удлиненные, шарообразные, звездчатые и т. д. Несмотря на такое разнообразие, все клетки растений можно объединить в две группы: паренхимные, если их размеры во всех направлениях примерно одинаковы, и прозенхимные, если длина клеток намного превышает их ширину. Величина клеток также разнообразна и колеблется от нескольких микрон до нескольких десятков сантиметров.
Растительная клетка у большинства растений с наружной стороны покрыта оболочкой, которую часто называют клеточной стенкой, особенно в тех случаях, когда речь идет о клетках, соединенных между собой. Под оболочкой располагается живое содержимое, или протопласт. Клетки без оболочки называются голыми. Они встречаются у некоторых низших растений. Клетки жизнедеятельны сравнительно ограниченное время и в процессе существования организма постоянно обновляются. Однако в ряде случаев после отмирания протопласта клетка не погибает, поскольку ее оболочка берет на себя некоторые функции.
Оболочка у молодых клеток состоит из целлюлозы, гемицеллюлозы и пектиновых веществ. По мере же роста и специализации клеток она часто пропитывается такими веществами, как лигнин, суберин, кутин, или инкрустируется солями кремниевой кислоты. Это вызывает соответственно одревеснение, опробковение, кутинизацию, минерализацию.
В многоклеточном организме клетки либо плотно прилегают одна к другой, либо между ними имеются промежутки, так называемые межклетники, обычно заполненные воздухом.

| Немембранные (рибосомы, микротрубочки, микрофиламенты) | 1-мембранные (ЭПР, аппарат Гольджи, плазмалемма, вакуоли, лизосомы, пероксисомы) | 2-мембранные (митохондрии, пластиды) |
Рис. 2. Общий план строения растительной клетки
В типичном случае растительная клетка состоит из протопласта (живого содержимого) и окружающей его оболочки – клеточной стенки.
Протопласт представляет собой цитоплазму с заключенными в нее органоидами (ядро, пластиды, митохондрии, комплекс Гольджи, сферосомы, рибосомы, эндоплазматический ретикулум, лизосомы). Количество органоидов и их состав зависят от таксономического положения растения, специфики жизнедеятельности клетки и от ее возраста.
Протопласт можно подразделить на цитоплазму и ядро. Цитоплазма состоит из гиалоплазмы и органелл. Гиалоплазма представляет собой непрерывную водную коллоидную фазу клетки и обладает определенной вязкостью. Она способна к активному движению за счет трансформации химической энергии в механическую. Гиалоплазма связывает все находящиеся в ней органеллы, обеспечивая их постоянное взаимодействие. Через гиалоплазму идет транспорт аминокислот, жирных кислот, нуклеотидов, сахаров, неорганических ионов, перенос АТФ.
Часть структурных белковых компонентов гиалоплазмы формирует надмолекулярные агрегаты – микротрубочки и микрофиламенты, которые образуют цитоскелет клетки. Микротрубочки представляют собой полые цилиндры диаметром 25 нм и располагаются параллельно друг другу близко к плазмалемме. Они образуют сеть интерфазных клеток, волокна веретена деления, входят в состав жгутиков. Предполагается, что они участвуют в поддержании формы протопласта, во внутриклеточном транспорте, перемещении органелл, ориентации образуемых плазмалеммой микрофибрилл целлюлозы.
Микрофиламенты в диаметре составляют 4–10 нм и состоят из спирально расположенных белковых субъединиц. Они являются сократимыми элементами цитоскелета и предположительно участвуют в движении цитоплазмы и перемещении органелл.
Органеллы – это структурно-функциональные единицы цитоплазмы. В клетке выделяют 3 типа органелл: немембранные, одномембранные и двумембранные.
Рибосомы относят к немембранным. Они состоят из рибонуклеопротеидов, включающих большую и малую субъединицы. Предшественники рибосом образуются в результате деятельности ядрышка, а окончательное формирование рибосом происходит в цитоплазме. Рибосомы могут располагаться в гиалоплазме, на мембранах гранулярного ЭПР, в митохондриях и пластидах. Каждая клетка содержит десятки тысяч или миллионы рибосом. Их основная функция – синтез белка. На поверхности гранулярного ЭПР рибосомы образуют полисомы, прикрепляясь к одной молекуле и-РНК, несущей информацию о первичной структуре белка.
К одномембранным органеллам относятся плазмалемма, эндоплазматический ретикулум, аппарат Гольджи, лизосомы, пероксисомы.
Плазмалемма является наружной поверхностной мембраной цитоплазмы. Она плотно прилегает к оболочке клетки и отделяет протопласт от окружающей среды. Плазмалемма хорошо проницаема для воды, которая проникает в клетку путем диффузии. Для крупных молекул она обычно непроницаема (барьерная функция). Мелкие молекулы и ионы проходят через плазмалемму с разной скоростью, поскольку она ограничивает их свободную диффузию и часто осуществляет перенос (транспортная функция). Кроме того, она выполняет функции синтеза микрофибрилл целлюлозы клеточной оболочки и восприятия гормональных сигналов.
Эндоплазматический ретикулум (ЭПР) представляет собой систему субмикроскопических канальцев, пронизывающих гиалоплазму, соединенных друг с другом и ограниченных одинарной мембраной. Различают два типа ЭПР: гранулярный и агранулярный. Гранулярный (шероховатый, г-ЭПР) несет на своих мембранах прикрепленные рибосомы. Состоит из канальцев и уплощенных цистерн. Цистерны ЭПР продырявлены многочисленными «окнами». Количество цистерн г-ЭПР колеблется в зависимости от типа клетки и стадии ее развития. Г-ЭПР выполняет функции синтеза белков (на прикрепленных к мембране снаружи полисомах), направленного транспорта макромолекул и ионов в клетке, участвует в образовании вакуолей, лизосом, диктиосом, а также взаимосвязывает все структурно-функциональные единицы клетки.
Агранулярный ЭПР (гладкий, а-ЭПР) лишен рибосом, имеет вид узких трубочек, пузырьков, цистерн. Обычно он развит слабее, чем г-ЭПР. Основная функция – синтез липофильных веществ и их транспорт. Хорошо развит в клетках, синтезирующих эфирные масла, смолы, каучук.
Аппарат Гольджи состоит из диктиосом и пузырьков. Диктиосома представляет собой стопку из 5–7 (до 20) дисковидных цистерн диаметром 1 мкм, ограниченных мембраной. На поперечном срезе диктиосомы цистерны имеют вид парных мембран, прямых или дугообразно согнутых. По краям диктиосомы переходят в систему тонких ветвящихся трубочек. Они имеют регенерационный полюс, на котором формируются цистерны из мембран ЭПР, и секреторный полюс, где отчленяются пузырьки Гольджи. В цистернах происходит синтез и транспорт аморфных полисахаридов, прежде всего – гемицеллюлоз и пектиновых веществ матрикса клеточной оболочки. Кроме того, аппарат Гольджи принимает участие в обновлении плазмалеммы, транспорте белков, образовании вакуолей и лизосом.
Лизосомы представляют собой мелкие цитоплазматические вакуоли и пузырьки – производные ЭПР или аппарата Гольджи. Они содержат гидролитические ферменты и выполняют функцию разрушения (переваривания) отдельных участков цитоплазмы собственной клетки, которое заканчивается образованием на ее месте цитоплазматической вакуоли.
Пероксисомы (микротельца) – мелкие пузырьки, в которых происходит процесс фотодыхания (окисления продуктов фотосинтеза кислородом). Образующийся при этом пероксид водорода, токсичный для клетки, разлагается под действием фермента каталазы.
К двумембранным органеллам относят митохондрии и пластиды.
Митохондрии у животных и растений имеют сходное строение. Снаружи они ограничены оболочкой, состоящей из двух мембран и светлого промежутка между ними. Наружная мембрана контролирует обмен веществ между митохондрией и гиалоплазмой. Внутренняя образует многочисленные выросты в полости митохондрии, называемые кристами. Пространство между кристами заполнено матриксом, в котором встречаются рибосомы, нити ДНК. Основная функция митохондрий – синтез АТФ из АДФ, т. е. обеспечение энергетических потребностей клетки. Синтез АТФ идет за счет окисления сахаров и называется окислительным фосфорилированием. Этот процесс протекает на внутренней мембране митохондрий, активная поверхность которой многократно увеличена за счет образования большого количества крист. Кроме того, митохондрии способны к синтезу белков, происходящему на их собственных рибосомах под контролем митохондриальной ДНК. В некоторых клетках митохондрии участвуют в синтезе липидов, углеводородов.
Пластиды встречаются только в растительных клетках. Выделяют 3 типа пластид: хлоро-, лейко- и хромопласты. Они отличаются друг от друга составом пигментов (цветом), строением и выполняемыми функциями.
Хлоропласты имеют зеленый цвети встречаются во всех зеленых органах растения (листьях, стеблях, незрелых плодах). Они содержат зеленый пигмент хлорофилл, который находится в хлоропластах в нескольких формах. Кроме хлорофилла в них содержатся пигменты, относящиеся к группе каротиноидов, в частности желтый (ксантофилл) и оранжевый (каротин), но обычно они маскируются хлорофиллом.
Хлоропласты, как правило, имеют линзовидную форму и сложное строение. Снаружи они ограничены оболочкой, состоящей из двух мембран. У хлоропластов, особенно высших растений, значительно развиты внутренние мембранные поверхности, имеющие форму плоских мешочков, называемых тилакоидами (ламеллами). На их мембранах находится хлорофилл. Тилакоиды могут располагаться одиночно, но чаще собраны в стопочки – граны. Внутренняя среда пластид называется стромой. В строме хлоропластов всегда встречаются пластоглобулы (включения жирных масел, в которых растворены каротиноиды), а также рибосомы, светлые зоны с нитями ДНК, а в некоторых случаях – крахмальные зерна, белковые кристаллы. Основная функция хлоропластов – фотосинтез. Кроме того, в них, как и в митохондриях, происходит процесс образования АТФ из АДФ, который называется фотофосфорилированием. Хлоропласты способны также к синтезу и разрушению полисахаридов (крахмала), некоторых липидов, аминокислот, собственного белка.
Лейкопласты – бесцветные, мелкие пластиды, встречающиеся в запасающих органах растений (клубнях, корневищах, семенах и т. д. ). Для лейкопластов характерно слабое развитие внутренней системы мембран, представленной одиночными тилакоидами, иногда трубочками и пузырьками. Остальные компоненты лейкопластов (оболочка, строма, рибосомы, ДНК, пластоглобулы) сходны с описанными для хлоропластов. Основная функция лейкопластов – синтез и накопление запасных питательных веществ, в первую очередь крахмала, иногда белков. Лейкопласты, накапливающие крахмал, называют амилопластами, белок – протеопластами, жирные масла – олеопластами.
Пластиды, окрашенные в желтые, оранжевые, красные цвета, носят название хромопластов. Их можно встретить в лепестках (лютик, одуванчик, тюльпан), корнеплодах (морковь), зрелых плодах (томат, роза, рябина, хурма) и осенних листьях. Яркий цвет хромопластов обусловлен наличием каротиноидов, растворенных в пластоглобулах. Внутренняя система мембран в данном типе пластид, как правило, отсутствует. Хромопласты имеют косвенное биологическое значение: яркая окраска лепестков и плодов привлекает опылителей и распространителей плодов.
Вакуоли содержатся почти во всех растительных клетках. Они представляют собой полости, заполненные клеточным соком и ограниченные от цитоплазмы мембраной – тонопластом. Для большинства зрелых клеток растений характерна центральная вакуоль. Она, как правило, настолько крупна (70–90 % объема клетки), что протопласт со всеми органеллами располагается в виде очень тонкого постенного слоя. Клеточный сок, содержащийся в вакуоли, представляет собой водный раствор различных веществ, являющихся продуктами жизнедеятельности протопласта. В его состав могут входить углеводы (сахара и полисахариды), белки, органические кислоты и их соли, минеральные ионы, алкалоиды, гликозиды, танины и другие растворимые в воде соединения.
Вакуоли в растительных клетках выполняют две основные функции: накопление запасных веществ, отходов и поддержание тургора. На второй функции остановимся подробнее. Концентрация ионов и сахаров в клеточном соке вакуоли, как правило, выше, чем в оболочке клетки. Поэтому при достаточном насыщении оболочки водой последняя будет поступать в вакуоль путем диффузии. Такой однонаправленный транспорт воды через полупроницаемую мембрану носит название «осмос». Поступающая в клеточный сок вода оказывает давление на постенный протопласт, а через него – на оболочку, вызывая напряженное, упругое ее состояние, или тургор. Он дает сочным органам растения форму и положение в пространстве и является одним из факторов роста клетки.
Если клетку поместить в гипертонический раствор осмотически активного вещества (NaCl, KNO3, сахарозы), т. е. в раствор с большей концентрацией, чем концентрация клеточного сока, то начнется осмотический выход воды из вакуоли. В результате этого объем ее сокращается, протопласт отходит от оболочки по направлению к центру клетки, тургор исчезает. Это явление обратимо и носит название «плазмолиз».
Клеточная оболочка – структурное образование на периферии клетки, придающее ей прочность, сохраняющее ее форму и защищающее протопласт. Оболочка, как правило, бесцветна и прозрачна, легко пропускает солнечный свет. По ней могут передвигаться вода и растворенные низкомолекулярные вещества. Оболочки соседних клеток соединены между собой пектиновыми веществами, образующими срединную пластинку.
Скелетным веществом оболочки клеток высших растений является целлюлоза. Молекулы целлюлозы, представляющие собой очень длинные цепи, собраны по нескольку десятков в группы – микрофибриллы. В них молекулы располагаются параллельно друг другу и сшиты многочисленными водородными связями. Они обладают эластичностью, высокой прочностью и создают структурный каркас оболочки. Микрофибриллы целлюлозы погружены в аморфный матрикс оболочки, состоящий в основном из гемицеллюлоз и пектиновых веществ. Молекулы матричных полисахаридов значительно короче молекул целлюлозы. Их цепи располагаются в оболочке достаточно упорядоченно и образуют многочисленные поперечные (ковалентные) связи как друг с другом, так и с целлюлозными микрофибриллами. Эти связи значительно повышают прочность клеточной оболочки. В зависимости от типа ткани, в состав которой входит клетка, в матриксе оболочки могут быть и другие органические (лигнин, кутин, суберин, воск) и неорганические (кремнезем, оксалат кальция) вещества.
В образовании структурных элементов клеточной оболочки принимают участие плазмалемма, аппарат Гольджи и микротрубочки. На плазмалемме происходит синтез микрофибрилл целлюлозы, а микротрубочки способствуют их ориентации. Аппарат Гольджи выполняет функцию образования веществ матрикса, в частности гемицеллюлоз и пектиновых веществ.
Различают первичную и вторичную клеточные оболочки. Меристематические и молодые растущие клетки, реже клетки постоянных тканей, имеют первичную оболочку, тонкую, богатую пектином и гемицеллюлозой. Вторичная клеточная оболочка образуется по достижении клеткой окончательного размера и накладывается слоями на первичную со стороны протопласта. Она обычно трехслойная, с большим содержанием целлюлозы. Для многих клеток (сосуды, трахеиды, механические волокна, клетки пробки) образование вторичной оболочки является основным моментом их высокоспециализированной дифференциации. Протопласт клетки при этом отмирает, и основную функцию клетки выполняют за счет мощной вторичной оболочки.
Плазмодесмы присущи только растительным клеткам. Они представляют собой тонкие цитоплазматические тяжи, соединяющие соседние клетки. В одной клетке может содержаться от нескольких сотен до десятков тысяч плазмодесм. Стенки канала плазмодесмы выстланы плазмалеммой, непрерывной по отношению к плазмалемме смежных клеток. По центру канала проходит мембранный цилиндр – центральный стержень плазмодесмы, соединенный с мембраной ЭПР. Между центральным стержнем и плазмалеммой в канале находится гиалоплазма. Плазмодесмы выполняют функцию межклеточного транспорта веществ.
Порами называют неутолщенные места оболочки (углубления), лишенные вторичной оболочки. Они содержат тончайшие отверстия, через которые проходят плазмодесмы. По форме порового канала различают простые и окаймленные поры. У простых диаметр канала приблизительно одинаков на всем протяжении от полости клетки до первичной оболочки и канал имеет форму узкого цилиндра. У окаймленных (трахеальные элементы) канал суживается в процессе отложения вторичной оболочки; поэтому внутреннее отверстие поры, выходящее в полость клетки, значительно уже, чем наружное, упирающееся в первичную оболочку. В смежных клетках поры располагаются напротив друг друга. Это облегчает транспорт воды и растворенных веществ от клетки к клетке. Общие поры имеют вид канала, разделенного перегородкой из срединной пластинки и первичной оболочки (замыкающая пленка поры).
Образование включений в растительных клетках связано с локальной концентрацией некоторых продуктов обмена веществ в определенных ее участках: в гиалоплазме, различных органеллах, реже в клеточной оболочке. Такое избыточное накопление веществ часто приводит к выпадению их в осадок в аморфном виде или в форме кристаллов. В функциональном отношении включения представляют собой временно выведенные из обмена веществ клетки соединения (запасные вещества) или конечные продукты обмена. К первой категории включений относят крахмальные зерна, липидные капли и отложения белков; ко второй – кристаллы некоторых веществ.
Крахмальные зерна – наиболее распространенные и важные включения растительных клеток, образующиеся только в строме пластид живых клеток. В хлоропластах на свету откладываются зерна ассимиляционного (первичного) крахмала, образующиеся при избытке продуктов фотосинтеза – сахаров. Образование осмотически неактивного крахмала предотвращает вредное повышение осмотического давления в фотосинтезирующих клетках.
Ночью, когда фотосинтеза нет, ассимиляционный крахмал с помощью ферментов гидролизуется до сахаров и транспортируется в другие части растения. Значительно большего объема достигают зерна запасного (вторичного) крахмала, откладывающиеся в лейкопластах (амилопластах).
Различают простые, полусложные и сложные зерна вторичного крахмала. Если в амилопласте имеется один центр образования, вокруг которого откладываются слои крахмала, то образуется простое зерно, если два или более – сложное зерно, состоящее из нескольких простых. Полусложное зерно образуется в том случае, если крахмал сначала откладывается вокруг нескольких центров, а затем после соприкосновения простых зерен вокруг них возникают общие слои. Видимая слоистость крахмальных зерен обусловлена неодинаковым обводнением слоев крахмала.
Крахмал является основным запасным веществом растений и может накапливаться в специализированных органах: корневищах, клубнях, семенах, плодах и т. д. Каждому виду растений свойственна определенная форма крахмальных зерен.
Липидные капли встречаются практически во всех растительных клетках, хотя численность и размеры их колеблются. Наиболее богаты ими семена и плоды, в которых они могут быть преобладающим по объему компонентом протопласта. Липидные капли, как правило, накапливаются непосредственно в гиалоплазме. Они представляют собой сферические тела обычно субмикроскопического размера. В световом микроскопе видны как сильнопреломляющие свет точки.
Белковые включения содержатся в различных частях клетки: в нуклеоплазме и перинуклеарном пространстве ядра, гиалоплазме, строме лейкопластов, вакуолях, цистернах ЭПР, матриксе митохондрий. Запасные белки чаще всего откладываются в виде зерен округлой или овальной формы, называемых алейроновыми. Если алейроновые зерна не имеют заметной внутренней структуры, их называют простыми. Иногда же в них среди аморфного белка можно заметить один или несколько белковых кристаллов. В отличие от настоящих кристаллов кристаллы белка набухают в воде, слабых кислотах и щелочах, окрашиваются красителями; поэтому их называют кристаллитами. Кроме того, в этих зернах встречаются блестящие бесцветные тельца округлой формы — глобоиды. Алейроновые зерна, содержащие кристаллиты и глобоиды, называют сложными. При обогащении клетки водой такие зерна растворяются. Алейроновые зерна разных видов растений, подобно крахмальным, имеют определенную структуру.
В растительных клетках часто встречаются кристаллы оксалата кальция, представляющие собой конечные продукты обмена. Они откладываются обычно в вакуолях. Форма этих кристаллов довольно разнообразна и часто специфична для определенных групп растений. Это могут быть одиночные кристаллы ромбоэдрической, октаэдрической или удлиненной формы (клетки наружных отмерших чешуек луковиц лука), друзы – шаровидные образования, состоящие из многих мелких сросшихся кристаллов (клетки корневищ, коры, корки, черешков и эпидермы листьев многих растений), рафиды – кристаллы в виде пучков игл (стебель и листья винограда), кристаллический песок – скопления множества мелких одиночных кристаллов (паренхимные клетки многих пасленовых, бузины). Наиболее часто встречающаяся форма кристаллов – друзы.
Кроме оксалата кальция кристаллические включения иногда состоят из карбоната кальция или кремнезема. Последний откладывается в клеточной оболочке.
Ядро представляет собой обязательный органоид живой клетки. Оно всегда располагается в цитоплазме. В молодой клетке обычно занимает центральное положение. При дифференциации клетки и образовании в ней большой центральной вакуоли отодвигается к клеточной оболочке, но всегда отделено от нее слоем цитоплазмы. Иногда ядро остается в центре клетки, окруженным цитоплазмой (т. н. ядерный кармашек). Цитоплазма при этом связана с постенным слоем тонкими тяжами.
Ядро отделено от цитоплазмы двухмембранной ядерной оболочкой, пронизанной многочисленными порами. Содержимое интерфазного (неделящегося) ядра составляют нуклеоплазма и погруженные в нее оформленные элементы – ядрышки и хроматин.
Ядрышки – сферические, довольно плотные тельца, состоящие из рибосомальной РНК, белков и небольшого количества ДНК. В ядре содержатся одно-два, иногда несколько ядрышек. Их основная функция – синтез р-РНК и образование рибонуклеопротеидов (рРНК+белок)-предшественников рибосом. Предрибосомы из ядрышка попадают в нуклеоплазму и через поры в ядерной оболочке переходят в цитоплазму, где и заканчивается их формирование.
Хроматин содержит почти всю ДНК ядра. В интерфазном ядре он имеет вид длинных тонких нитей, представляющих собой двойную спираль ДНК, закрученных в виде рыхлых спиралей более высокого порядка (суперспиралей). ДНК связана с белками-гистонами, располагающимися, подобно бусинкам, на нити ДНК. Хроматин, будучи местом синтеза различных РНК (транскрипции), представляет собой особое состояние хромосом, выявляющихся при делении ядра. Можно сказать, что хроматин – это функционирующая, активная форма хромосом. Дело в том, что в интерфазном ядре хромосомы сильно разрыхлены и имеют большую активную поверхность. Такое диффузное распределение генетического материала наилучшим образом соответствует контролирующей роли хромосом в обмене веществ клетки. Следовательно, хромосомы присутствуют в ядре всегда, но в интерфазной клетке не видны, потому что находятся в деконденсированном (разрыхленном) состоянии.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|