- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Нарушения метаболизма при СД
103. Взаимосвязь обмена углеводов, жиров и белков. Пункты взаимосвязей метаболизма глюкозы и липидов, углеводов и аминокислот, аминокислот и липидов. Значение в этих процессах цикла трикарбоновых кислот, апотомического окисления глюкозы, глюконеогенеза.
Обмен веществ - как единое целое , все биохимические процессы тесно взаимосвязаны. Обмен белков, жиров и углеводов представляют собой единое целое.
Эта взаимосвязь осуществляется через, так называемые узловые, метаболиты- это общие звенья превращений, к ним относятся: глюкозо-6-фосфат(в гликолизе),пируват(в гликолизе) глицерол-3-фосфат(в обмене углеводов и липидов), Ацетил-КоА(Цикл Кребса) , альфа-кетоглутарат(цикл кребса), оксалоацетат и тд
Основные метаболические процессы, которые объединяют обмен веществ: гликолиз,глюконеогенез, переаминирование, цикл Кребса и тд
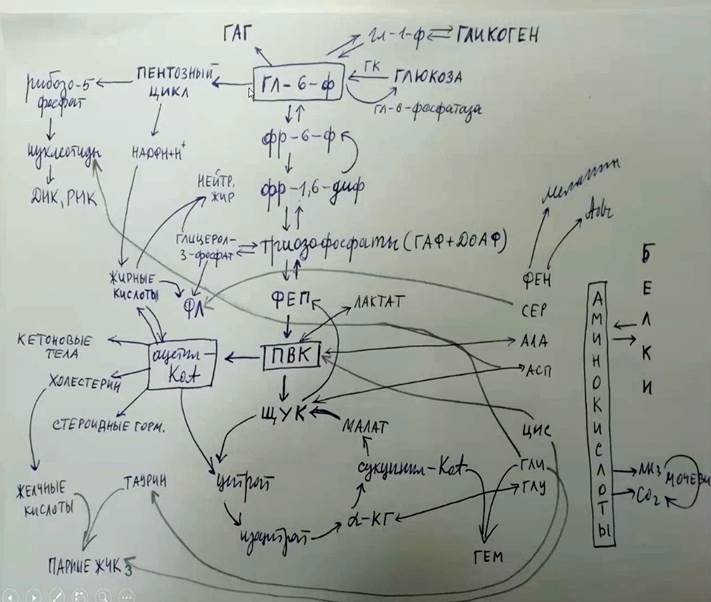
ОПИСАНИЕ
Сначала мы должны изобразить обмен углеводов(сверху пишем ключевой метаболит углеводного обмена-глюкозо-6-фосфат в рамочке)
Направо глюкоза (изогнутая стрелочка) фермент глюкозо-6-фосфатаза-это обратная реакция, и прямая реакция-фермент ГК(гексокиназа)
Также гл-6-ф переходит в → гл-1-ф и в гликоген(ферм-гликогенсинтетаза)
Налево наверх ГАГ(гликозоаминогликаны)
Путь превращения глюкозы-это пентозныйцикл(стрелочка налево)-продукт окислительный фазы НАДФН+Н(стрелочка вниз);далее от пентозного цикла – рибозо-5-фосфат(используется для синтеза нуклеотидов днк и рнк)
Вниз стрелочка от гл-6-ф- основные метаболиты гликолиза(фр-6-фосфат →фр-1.6-дифосфат прямая и обратная реакции →триозофосфаты(ГАФ(глицеральдегидфосфат) и ДОАФ (диоксиацетонфосфат) )(обратимая реакция) →фосфоенолпируват(ФЕП),который превращается в пируват-так как это анаэробный процесс-гликолиз-направо уводим стрелочку →лактат(последний продукт гликолиза) обратимый процесс(ферм-ЛДГ)
Далее вниз идет цикл кребса. ПВК дает ацетил коа(стрелочка влево)-это ключевой метаболит липидного обмена(обратный процесс невозможен!!!
ПВК(стрелочка вниз) → ЩУК(также обратный процесс ЩУК → ФЕП →триозофосфаты →фр1.6диф →фр-6-ф → гл-6-ф глюкоза --- это и есть глюконеогенез)
Ацетил КоА вместе с ЩУК дает цитрат, дает стероидные гормоны холестерин и кетоновые тела(обратимо), жирные кислоты(обратимо)
Жирные кислоты(наверх стрелочка) используется для синтеза нейтрального жира и для этого нужен НАДФН+Н и для синтеза нейтрального жира нужен глицерол-3-фосфат(напротив триозофосфатов,триозофосфаты, гаф+доаф являются источниками глицерол-3-фосфата)
Кроме того, жирные кислоты вместе с глицерол-3-фосфатом дают фосфолипиды
*в центре и сверху схемы обмен углеводов, слева обмен липидов, справа обмен белков
Далее, ЩУК → цитрат →изоцитрат →альфа кетоглутарат →сукцинил-коа →малат →ЩУК – это цикл трикарбоновых кислот(в сокр варианте)
Справа рисуем белки → они превращаются в аминокислоты(показываем только основные)
1)фениаланин →дает меланин и адреналин
2)серин → к фосфолипидам, то есть жирные кислоты , глицерол-3-фосфат и серин дают фосфолипиды(например фосфотидилсерин)
3)аланин- через реакцию переаминирования превращается в ПВК (ферм АЛТ, обратимая)
4)аспартат → ЩУК (ферм АСТ, обратимая); → к нуклеотидам(используется для пуриновых и пиримидиновых оснований)
5) цистеин → в ПВК; → к таурину,с которым образует парные желчные кислоты(например тауровая)
6)глицин-вместе с сукцинил-КоА дают ГЕМ ; → также дает парные желчные кислоты(например гликохолевая кислота) ; также еще глицин встраивается в пуриновое кольцо(т.е используется для синтеза нуклеотидов)
7)глутамат – превращается в процессе переаминирования в альфа-кетоглутарат
При распаде аминокислот → аммиак, углекислый газ → цикл мочевины
Таким образом, все виды метаболизма тесно между собой связаны, но обратный процесс превращения Ацетил-КоА в ПВК невозможен(!!) то есть белки и углеводы легко превращаются в липиды, а обратный процесс практически невозможен.
Всего у нас в организме происходит 5000 биохимических реакций – реактом(совокупность всех реакций в орагнизме)
Это основные метаболические превращения и их взаимосвязи.

Березов
В настоящее время экспериментально обосновано существование четырех главных этапов распада молекул углеводов, белков и жиров, которые интегрируют образование энергии из основных пищевых источников. На I этапе полисахариды расщепляются до моносахаридов (обычно гексоз); жиры распадаются на глицерин и высшие жирные кислоты, а белки – на составляющие их свободные аминокислоты. Следует подчеркнуть, что указанные процессы в основном являются гидролитическими, поэтому освобождающаяся в небольшом количестве энергия почти целиком используется организмами в качестве тепла. На II этапе мономерные молекулы (гексозы, глицерин, жирные кислоты и аминокислоты) подвергаются дальнейшему распаду, в процессе которого образуются богатые энергией фосфатные соединения и ацетилКоА. В частности, при гликолизе гексозы расщепляются до пировиноградной кислоты и далее до ацетил-КоА. Этот процесс сопровождается образованием ограниченного числа богатых энергией фосфатных связей путем субстратного фосфорилирования. На этом этапе высшие жирные кислоты аналогично распадаются до ацетил-КоА, в то время как глицерин окисляется по гликолитическому пути до пировиноградной кислоты и далее до ацетил-КоА. Для аминокислот ситуация на II этапе несколько отлична. При преимущественном использовании аминокислот в качестве источника энергии (при дефиците углеводов или при сахарном диабете) некоторые из них непосредственно превращаются в метаболиты лимоннокислого цикла (глутамат, аспартат), другие – опосредованно через глутамат (пролин, гистидин, аргинин), третьи – в пируват и далее в ацетил-КоА (аланин, серин, глицин, цистеин). Наконец, ряд аминокислот, в частности лейцин, изолейцин, расщепляется до ацетил-КоА, а из фенилаланина и тирозина, помимо ацетил-КоА, образуется оксалоацетат через фумаровую кислоту. Как видно, II этап можно назвать этапом образования ацетил-КоА, являющегося по существу единым (общим) промежуточным продуктом катаболизма основных пищевых веществ в клетках. На III этапе ацетил-КоА (и некоторые другие метаболиты, например α-кетоглутарат, оксалоацетат) подвергаются окислению («сгоранию») в цикле ди- и трикарбоновых кислот Кребса. Окисление сопровождается образованием восстановленных форм НАДН + Н+ и ФАДН2 . На IV этапе осуществляется перенос электронов от восстановленных нуклеотидов на кислород (через дыхательную цепь). Он сопровождается образованием конечного продукта – молекулы воды. Этот транспорт электронов сопряжен с синтезом АТФ в процессе окислительного фосфорилирования.
104. Сахарный диабет. Типы, причины возникновения и основные проявления. Нарушения метаболизма при сахарном диабете, обмена углеводов, липидов, аминокислот. Механизмы возникновения кетонемии, кетонурии, гиперхолестеринемии, гипергликемии и других нарушений при сд. Гликозилированные белки. Их значение при возникновении ангиопатии, сахарные кривые в диагностике диабета.
Сахарный диабет- заболевание, связанное с нарушением всех видов обменов веществ, но главным образом метаболизмом глюкозы на фоне недостаточности инсулина, и характеризуется это заболевание гипергликемией, глюкозурией и развитием осложнений с повреждением сосудов и нервных путей.
Типы СД: 1) СД 1-го типа (инсулинзависимый) – для него характерен абсолютный дефицит инсулина. Как правило это заболевание аутоиммунное, при котором образуются ауто-антитела к бета-клеткам(т.е повреждаются бета-клетки, инсулин не синтезируется)
-болеют молодые люди до 30 лет, дети; до 20% всех случаев СД.
2) СД 2-го типа (инсулиннезависимый) – связан с нарушением рецепции инсулина, характерна инсулинорезистентность(т.е инсулин есть, но он не рецептируется и возникает нечувствительность к инсулину
-страдают люди старше 40 лет на фоне ожирения. Более 80% всех случаев СД.
Нарушения метаболизма при СД
Патология углеводного обмена
1) Нарушается поступление глюкозы в клетку- это связано с тем, что не синтезируется ГЛЮТ-4(это транспортный белок)-глюкозы не поступает. Страдают мышцы, жировая ткань.
2)Нарушается использование глюкозы в тканях и клетках- опять таки из-за дефицита инсулина не синтезируются ключевые ферменты метаболизма углеводов(гексокиназы, фосфофруктокиназы, глюкозо-6-фосфатдегидрогеназы, гликогенсинтетазы и др)
3) Происходит активация глюконеогенеза
4) Снижается синтез глюкозоаминогликанов, гиалуроновой кислоты, нарушается барьерная функция кожи
Патология липидного обмена
1) Если глюкоза в клетки не поступает, происходит усиление использования липидов на энергетические цели
2) Вследствие происходит активация липолиза, повышается концентрация Ацетил-КоА(ацетил-коа превращается в кетоновые тела)
3)Повышение синтеза кетоновых тел- поэтому развивается характерный признак СД-это кетонурия и кетонемия
4)Повышается синтез холестерина- у таких пациентов склонность к атеросклерозу
5)Активируется синтез глюко- и минералокортикоидов, образуются стероидные горомны- и она стимулируют глюконеогенез и нарушение водно-солевого обмена.
Патология обмена белков
1)Поскольку снижается роль глюкозы, повышается роль белков, происходит активация катаболизма и аминокислоты используются для глюконеогенеза
2)увеличивается синтез мочевины, поскольку много образуется аммиака – развивается отрицательный азотистый баланс
3)угнетается синтез белка, синтез антител, возникает иммунодефицит, нарушается заживление ран
Патология водно-солевого обмена
1) глюкозурия- глюкоза появляется в моче, развивается при превышении уровня глюкозы крови 1,6-1,8 г/, то когда она превышает порог она появляется в моче, в норме быть в моче не должно
2)полиурия- когда у пациента выделяется более 3 л мочи в сутки
3) полидипсия- неутолимая жажда
*диабет переводится как мочеизнурение
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|