- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Лекция 5.
Лекция 5.
ФЕНОМЕН ЖИЗНИ -MISTERIUM TREMENDUM[1]
Жизнь есть космическое явление, в чем-то резко отличное от косной материи.
X. Гюйгенс «Космотеорос»
1. ЧТО ТАКОЕ ЖИЗНЬ?
Удовлетворительного определения феномену жизни до сих пор не существует. При попытках выяснить, чем живое отличается от неживого, естественно, выделялись характерные свойства организмов, населяющих нашу планету, так как иные, неземные, формы жизни науке пока не известны. Понятие «жизнь» всегда отождествлялось с ее конкретной реализацией в условиях нашей планеты. Именно поэтому в большинстве учебников обычно идут по пути перечисления признаков, которые характерны для живого, но при рассмотрении этих признаков можно обнаружить, что практически все они встречаются и у объектов неживой природы. В таблице 1 приведен один из примеров такого сопоставления наиболее характерных признаков. Таким образом, попытки найти какой-либо абсолютный, характерный признак живого — занятие малоперспективное. В живых системах, как правило, не обнаруживается никаких свойств, которыми не обладали бы разные неживые, объекты.
Вместе с тем в пространстве рассмотренных в таблице 1 признаков, например, вирус или бактериофа вообще нельзя назвать живыми организмами в полном смысле слова. Скорее, это сложные композитные материалы, но решающие задачи поиска, узнавания и размножения путем уничтожения клетки или бактерии.
Чаще всего среди многочисленных признаков, отличающих, на первый взгляд, живое вещество от неживой материи, биологи выделяют следующие.
1. Способность быстро занимать (осваивать) все свободное пространство.
2.Движение — не только пассивное (под действием внешних сил), но и активное. Например, против течения воды, потоков воздуха, силы тяжести.
Таблица 1.
Наиболее характерные признаки живой и неживой материи
| № | Признаки живой материи | Признаки неживой материи |
| Характеризуется упорядоченной иерархической структурой | Все объекты неживой природы отвечают этому же условию и устроены по иерархическому принципу: элементарные частицы => атомы => молекулы => макромолекулы и т. д. | |
| Являются открытыми системами и получают энергию из окружающей среды, используя ее для поддержания своей высокой упорядоченности | Смерчи, тайфуны, ветер, молнии черпают энергию от Солнца; извержение вулканов, землетрясения, подвижка материков происходят за счет энергии из недр Земли. Таким образом, открытость живых систем — не специфический признак живого | |
| Способность реагировать на внешнее воздействие: (рецепция) — универсальное свойство всех живых систем | Намагничивание, электризация, свечение, поляризация, деформация, инерция, перемещение, разрушение и т. д. — это также реакция неживых объектов на внешние воздействия | |
| Способность запоминать информацию о предыдущих состояниях и адаптироваться к изменению внешних условий | В неживых объектах существуют проявления элементов памяти. Ответная реакция объектов неживой природы обычно направлена на «нейтрализацию» внешнего воздействия — это стремление сохранить свое исходное состояние (принцип Ле Шателье, закон Ленца, инерция Ньютона) | |
| Живые организмы изменяются и усложняются | Объекты в астрофизике (образование газопылевых облаков => туманностей => галактик), в геофизике (образование горячего ядра планет => сравнительно холодной мантии поверхности планет => тектонических плит => материков и океанов) и в химии (преобразование субстратов в продукты) также демонстрируют эволюционное изменение и усложнение | |
| Все живое размножается | Коацерватные капли органических веществ могут расти и делиться. Из растворов солей растут кристаллы. Кусочек, отломившийся от растущего кристалла, становится зародышем для подобного кристалла | |
| Живое способно к саморегуляции и регенерации повреждений | Устойчивые вихри, торнадо, ячейки Бенара — саморегулирующиеся системы. Кристаллы способны к регенерации дефектов (дислокаций). Саморегуляция и регенерация — характерна для неживой природы | |
| Живые объекты осуществляют обмен веществ с окружающей средой | Преобразование энергии — это свойство всей природы, а не специфическое свойство живых систем. Например, все реакции окисления (горения) обладают этим свойством | |
| Живые объекты обладают направленной подвижностью | Этим же свойством обладают ферромагнитные частицы в магнитном поле, ионы в электрическом поле, броуновские частицы в тепловом поле, частицы, имеющие массу, в гравитационном поле, тайфуны | |
| Живым объектам свойственна неравновесность состояния | Дожди, снегопады, лавины, водопады, ячейки Бенара, конвективные потоки и т. п. — это также неравновесные состояния |
Устойчивость при жизни (при весьма высокой физико-химической активности) и быстрое разложение после смерти.
Высокая приспосабливаемость (адаптация) к различным условиям внешней среды.
Феноменально высокая скорость протекания реакций. Она на несколько порядков выше, чем в неживом веществе.
Гусеницы некоторых насекомых, например, потребляют за день количество пищи, которое в 100-200 раз больше массы их тела. Дождевые черви за 150-200 лет пропускают через свои организмы слой почвы толщиной около метра. По представлениям В. И. Вернадского, практически все осадочные породы (слой до 3 км) на 95-99% переработаны живыми организмами.
Высокая скорость обновления живого вещества.
В среднем для биосферы она составляет 8 лет, для суши — 14 лет, для океана, где преобладают организмы с коротким периодом жизни — 33 дня.
При классификации по признаку живое — неживое часто оперируют еще двумя признаками.
Обмен вещества с внешней средой, характерный для живого вещества. Например, по А. И. Опарину: Живое вещество — сложные молекулярные агрегаты (белковые тела), характеризующиеся упорядоченным обменом веществ с внешней средой.
Отметим, что данное утверждение следует воспринимать как классификационный признак, но не как определение живого вещества. Во-первых, потому что он далеко не исчерпывает всей сущности живого вещества, во-вторых, в настоящее время установлено, что обмен веществ, хотя и в более простых формах, существует и в мире неживой природы.
Способность живой материи к тождественному воспроизводству.
Долгое время считалось, что это один из основных отличительных признаков, хотя и оставалось загадкой, каким образом в живых организмах осуществляется передача наследственных признаков от родителей к потомкам. Эта точка зрения среди биологов еще более утвердилась после 1950-х гг. в связи с исследованиями дезоксирибонуклеиновых кислот (ДНК), содержащихся в ядрах клеток живых организмов.
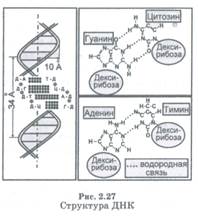 В 1953 г. английский биофизик Ф. Крик и его американский коллега Д. Уотсон, опираясь на результаты рентгеноструктурных исследований, расшифровали структуру молекулы ДНК и установили, что она состоит из двух винтообразных цепочек (содержащих десятки миллионов атомов каждая), свернутых вокруг друг друга в двойную спираль (рис. 2.27). Уотсон и Крик высказали предположение, что именно на этих цепочках «записана» вся генетическая информация, передаваемая от поколения к поколению, причем «запись» на одной из цепочек полностью продублирована на другой.
В 1953 г. английский биофизик Ф. Крик и его американский коллега Д. Уотсон, опираясь на результаты рентгеноструктурных исследований, расшифровали структуру молекулы ДНК и установили, что она состоит из двух винтообразных цепочек (содержащих десятки миллионов атомов каждая), свернутых вокруг друг друга в двойную спираль (рис. 2.27). Уотсон и Крик высказали предположение, что именно на этих цепочках «записана» вся генетическая информация, передаваемая от поколения к поколению, причем «запись» на одной из цепочек полностью продублирована на другой.
При делении клетки на две цепочки молекулы ДНК также разделяются. В обе новые клетки попадает по одной цепочке из каждой молекулы ДНК. Каждая новая клетка вдоль такой одиночной цепочки достраивает соответствующую ей вторую. Таким образом, двойные спирали ДНК оказываются восстановленными, а генетическая информация, записанная в них, — продублированной. Этот процесс получил название репликации (от лат. replicatio — повторение). В наборе молекул ДНК, находящемся в ядре любой клетки, полностью записана программа построения всего организма, в том числе и программа управления процессом репликации.
При всей значительности своих размеров молекулы ДНК состоят всего лишь из шести типов «строительных кирпичиков» (шести типов устойчивых групп атомов): дезоксирибозы, фосфорной кислоты и четырех органических азотистых оснований. Основу цепочек ДНК (ДФ-основу) составляют поочередно связанные друг с другом молекулы дезоксирибозы (Д) и фосфорной кислоты (Ф). К каждой молекуле дезоксирибозы присоединено также одно из азотистых оснований: аденин (А), тимин (Т), гуанин (Г) и цитозин (Ц). Сама по себе ДФ-основа еще не несет никакой информации, а служит лишь «подложкой» (или «строкой»), на которую информация записывается. Роль информативных символов для записи играют азотистые основания. В их последовательности заключена вся передаваемая из поколения в поколение информация.
Молекулу ДНК можно условно разделить на отдельные участки — гены, в каждом из которых записан один блок информации. Каждый ген отвечает за синтез в клетке того или иного вещества. Таким образом, молекулы ДНК через гены управляют всеми биохимическими реакциями, происходящими в организме, что и определяет, в конечном счете, все признаки этого организма. Две цепочки, образующие молекулу ДНК, являются в своем роде зеркальными. Цитозиновое и гуаниновое основания, входящие в разные цепочки, всегда находятся напротив друг друга. То же относится и к паре тимин — аденин. Процесс репликации молекул ДНК осуществляется весьма сложным образом, но суть его заключается в том, что у каждой из двух разделившихся цепочек выстраивается ее «зеркальное отображение».
Благодаря способности молекул ДНК к репликации процесс деления клеток формально может повторяться бесконечное количество раз. Поэтому разумно было бы считать, что настоящая жизнь на Земле возникла тогда, когда природа смогла создать способные к репликации носители наследственной информации — молекулы ДНК или им подобные.
Способность к торжественному воспроизводству, без сомнения, является важнейшим признаком живого вещества.
Однако оказалось, что способностью к воспроизводству себе подобных обладают и объекты неживой природы.
 |
Один из наиболее ярких классических примеров — кристаллизация и конденсация жидкостей. Для того чтобы при охлаждении пара его избыток выпал в виде росы, необходимы центры конденсации, а для образования кристаллической структуры, как правило, необходима кристаллическая «затравка» (фактически, по сравнению с живой природой, очень простой, но по функциям — аналог ДНК), структуру которой и пытаются повторить молекулы жидкости при кристаллизации. В реальной среде такие центры легко реализуются при наличии пыли и прочих микрочастиц в атмосфере. Однако если водяной пар специально очистить от посторонних взвесей, то при охлаждении пара его конденсация не происходит, так как пар «не знает», что ему делать. Такой пар называют переохлажденным. На этом принципе, в частности, работает камера Вильсона для регистрации заряженных элементарных частиц, невидимых глазом: если в объем с переохлажденным паром попадает такая частица, она выступает в качестве центра конденсации, и вдоль всего ее пути появляется след в виде сконденсировавшегося пара, аналогичный тому, который мы видим в небе, когда там достаточно высоко пролетает реактивный самолет.
Даже в домашних условиях можно поставить такой эксперимент. Известно, что гипосульфит плавится при температуре около 50°С. Если стакан расплавленного гипосульфита очистить от примесей, в том числе, от не расплавившихся взвешенных микрочастиц, то при медленном охлаждении, вплоть до комнатной температуры он будет оставаться жидким, как будто молекулы «не знают», что им можно построить кристаллическую структуру (рис. 2.28а, б). Однако если теперь в жидкий гипосульфит, находящийся при комнатной температуре (т.е. почти на 30°С ниже температуры кристаллизации), бросить кристаллик гипосульфита, практически сразу же начнется процесс кристаллизации. Уже через 5-7 мин. почти весь находящийся в стакане гипосульфит станет кристаллическим
(рис. 2.28б-г). Кристаллизация (самоорганизация) идет с выраженным выделением энергии (противодействие процессу кристаллизации) — температура повышается и может даже приблизиться к температуре плавления.
Существенно, что для кристаллизации всегда нужен образ структуры будущего кристалла, или, как говорят кристаллографы, — «затравка». Если характер электронных оболочек молекул и их энергетические возможности позволяют образовывать структуру «затравки», она реализуется. При этом кристалл будет стараться повторить даже дефекты (на языке биологии — мутации) структуры «затравки».
В связи с этим напомним, что для одних и тех же атомов и молекул, в зависимости от типа «затравки» и созданных внешних условий, могут реализовываться различные типы кристаллических структур. Углерод, например, может кристаллизоваться в структуры графита, каменного угля или алмаза. Вода также реализует несколько различающихся по структуре и физическим свойствам типов кристаллической структуры льда. Важно понимать, что структура кристалла зависит не только от сорта атомов, но и от созданных на данный момент внешних условий.
Конечно, возможностей для разнообразия при кристаллизации простых атомных структур гораздо меньше, чем при воспроизведении себе подобных в живой природе. Так, существует всего семь сингоний кристаллических решеток, из которых наиболее распространены всего 3-4. Поэтому в качестве «затравки» легко могут выступать и случайные примеси, в том числе другое вещество с близкой структурой. Например, железо при комнатной температуре обычно имеет объемно центрированную кубическую структуру. Однако если то же самое железо напылять на подложку из палладия, то оно кристаллизуется в виде гранецентрированной кубической структуры, повторяя структуру палладия, выступающего в данном случае в роли «затравки». Поэтому в реальной жизни кристаллизация и конденсация происходят практически всегда из-за наличия большого количества посторонних примесей и пыли в виде взвешенных частиц, и нам кажется, что все происходит само собой. Тем не менее, каждый кристалл несет в себе определенную информацию о своей «затравке», т. е. делает то же, что и ДНК в живой природе. Это позволяет, например, несмотря на, казалось бы, однотипное строение кристаллов алмаза, по анализу особенностей их кристаллической структуры и сопоставлению их с эталонными для различных месторождений, достаточно точно определять место добычи.
В 1950-е гг. химики научились синтезировать сложные высокомолекулярные соединения. В частности, было установлено, что длинные молекулярные цепочки, с химической точки зрения, повторяющие структуру белков и нуклеиновых кислот живой материи, могут образовываться в водных растворах под действием тепла и некоторых катализаторов. Таким образом, можно было предположить, что большие количества органических веществ могли образоваться в первичной земной атмосфере или даже еще в допланетном газопылевом облаке. Когда на поверхности Земли появилась жидкая вода, эти органические вещества растворились в ней, обретя подвижность, необходимую для зарождения жизни. Путем полимеризации «строительных кирпичиков», протекавшей под действием тепла и катализаторов, образовались различные высокомолекулярные соединения (в том числе, белково- и ДНК-подобные) и, в конечном счете, зародилась жизнь. Однако возникла не поддающаяся на сегодняшний день разумному объяснению проблема:
Между высокомолекулярными соединениями, которые удалось синтезировать в лабораториях, и реальными молекулами ДНК существует огромная разница.
В частности:
Принципиальные различия в записанной информации.
Современная наука пока не знает, каким образом произошла запись первичной нужной информации в молекуле ДНК.
Молекулы ДНК живой материи являются носителями огромного объема информации. Их энергетическое состояние соответствует вполне определенной совокупности команд, записанных в генах, по которым осуществляется согласованное строительство всего организма.
Энергетическое состояние же искусственно полученных ДНК, как правило, достаточно случайно. Вероятность случайной комбинации, которая образовала бы вполне осмысленный и согласованный набор команд исключительно мала[2].
Разовость возникновения жизни.
Возникновение жизни — это действительно феномен, особая точка в эволюции Вселенной, а именно — масштабная бифуркация.
С большой уверенностью можно говорить о том, что жизнь на Земле (а возможно, и во Вселенной в целом) возникла лишь единожды. Далее все живое порождалось только живым.
Факты, подтверждающие это:
• Глубочайшее биохимическое сходство всех существующих на Земле живых организмов.
В природе существует свыше 150 различных аминокислот, любая из которых, в принципе, могла бы участвовать в построении белково-подобных молекул. Однако белковые молекулы всех без исключения известных на Земле организмов состоят из соединенных в цепочки фрагментов одних и тех же 20 аминокислот. По-видимому, этот набор аминокислот просто соответствует выбранному природой случайно и именно разово при образовании первичных микроорганизмов, потомками которых является все, что ныне живет на нашей планете.
• Асимметрия живого вещества (гомохиральность строения). Хиральность — понятие, характеризующее зеркальную парность лево-симметричных и право-симметричных объектов в природе. Большинство веществ с упорядоченной структурой атомов (обычно в виде кристаллической решетки) хиральны, т. е. могут существовать в виде двух типов зеркально симметричных кристаллических решеток. Обычно в природе «левые» и «правые» разновидности кристаллов представлены в равной мере.
Молекулы аминокислот также имеют две симметричные формы структуры — левую и правую. В аминокислотах, синтезированных неорганическим путем левые и правые варианты молекул (изомеры) обычно смешаны в равных количествах. Поэтому следует ожидать, что на первичной Земле левые и правые формы аминокислот были представлены в равной мере.
 |
На этом фоне совершенно удивительным выглядит тот факт, что аминокислоты живых организмов представлены только левыми изомерами.
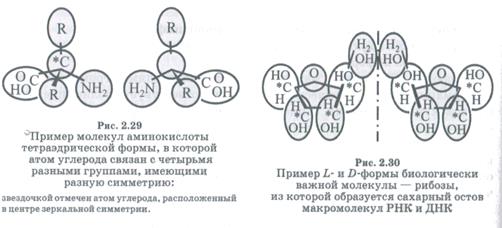
По какой-то причине природа не использовала при построении белковых молекул ни одной из правых форм аминокислот. Аминокислоты, которые входят в состав функциональных белков всех организмов, — это только L-аминокислоты (лат. Laevus — левый). Хотя короткие полипептиды с D-аминокислотами (лат. Dexter — правый) существуют (например, нейропептиды), но это не структурные, а сигнальные белки. Некоторые известные нам ферменты состоят более чем из 100 аминокислот, и все они находятся в L-форме. Если в активный центр фермента, построенного из L-аминокислот, включилась хотя бы одна D-аминокислота, то последний перестал бы функционировать. Организмы используют D-аминокислоту для информационных функций. С другой стороны, в РНК и ДНК при формировании остова используются только D-caxapa. На рисунках 2.29 и 2.30 приведены два примера хиральных соединений.
Проблема хиральности и способ спонтанного разделения — это не только чисто фундаментальная, но и практическая задача, например синтез хирально чистых наносоединений. Сегодня вся мировая фармакологическая промышленность переходит на хирально чистые лекарства. Это было инициировано известной историей, произошедшей в 1970-х гг. Тогда родилось несколько сотен детей с пороками развития костных тканей и отсутствием развитых конечностей — так называемой тюленьей болезнью. Причина состояла в том, что препарат одной из фармацевтических фирм, продаваемый как успокоительное и снотворное средство, содержал имидталоилглютаминовую кислоту в смешанном хиральном соединении. Раздельное испытание этих антиподов показало, что L-антипод вызывает уродства, тогда как D-антипод является безопасным. Это свидетельствует о том, насколько для биосистем важна гомохиральностъ.
Еще Л. Пастер, открывший левую симметрию живого вещества, отмечал, что она должна быть связана с действием в период возникновения жизни какого-то мощного асимметричного физического фактора. Если бы жизнь возникала постепенно в разное время и (или) в нескольких точках пространства независимо, то левые и правые формы аминокислот в живой материи должны были бы встречаться одинаково часто.
Схожесть генной структуры живых организмов. Результаты крупного международного научного проекта по расшифровке трех миллиардов нуклеотидных пар, содержащих генетическую информацию человека, завершенного в апреле 2003 г., показали, что генетический код всех организмов — единый, и различается лишь незначительно количеством генов.
Число обнаруженных генов в ДНК человека оказалось равным всего 30 000, а предполагалось, что их, по крайней мере, в три раза больше (порядка 100 000). Это число генов совпало с числом генов мыши, а качественно по геному мы отличаемся от мыши структурой всего 300 генов. Порядка 100 наших генов соответствуют геномам вирусов и бактерий. Правда, до сих пор непонятно, что представляют собой обширные нуклеотидные последовательности в промежутках между генами у человека и какую роль они играют в развитии организма.
2. КВАНТОВАЯ БИОЛОГИЯ ИЛИ ЭВОЛЮЦИЯ ПО ДАРВИНУ?
С позиций синергетики можно с уверенностью утверждать, что, хотя сама эволюция видов, описанная Дарвином, близка к реальной, механизм этой эволюции (эволюция по Дарвину) вряд ли состоятелен. Он предполагает плавное непрерывное (континуальное) изменение видов под действием внешних изменяющихся условий и постулирует, что движущими силами эволюции живого являются изменчивость, наследственность и естественный отбор. В ходе биологической эволюции на Земле образовались многие миллионы сменявших друг друга видов живых организмов.
Однако Ч. Дарвин ошибался, когда считал, что материалом, на основе которого действует естественный отбор, являются небольшие непрерывные, случайные изменения, обязательно встречающиеся даже в наиболее однородной популяции.
Для несложных систем (процессы кристаллизации, возможно, даже размножение вирусов) сценарий, предложенный Дарвином, в принципе, хотя и с некоторыми оговорками, может быть реализован на практике. Для основной же массы биообъектов наличие столь сложной структуры как клетка с цепочками ДНК, не позволяет реализовать механизм мутаций.
Именно с фактором отсутствия достаточной устойчивости структуры у простых самоорганизующихся систем типа вирусов объясняют сегодня трудности эффективной борьбы с ними при помощи медицинских препаратов, так как вирусы с помощью небольших мутаций легко модифицируются под их структуру. Новые, появившиеся после мутации, разновидности вирусов легко, без противоречий начинают сосуществовать с этими препаратами. Возможно только временное приостановление роста популяции вирусов на период, достаточный для создания достаточно крупной их колонии с видоизмененной структурой. Для сложных биообъектов такой вариант невозможен. Поэтому существует понятие яда, приводящего к гибели организма.
Для изменения вида должна произойти синхронная мутация многих систем жизнеобеспечения биообъекта, причем, не приводящая к их взаимному противодействию (отторжению). Вероятность такой одновременной синхронной мутации чрезвычайно мала. В случае если такое событие все же произойдет, то оно будет, скорее всего, напоминать скачок (бифуркацию), соответствующий существенному изменению вида (формально — появлению нового вида, а не плавному его изменению).
Открытием законов передачи последующим поколениям признаков (в том числе, рецессивных и доминантных), которыми различались родители, мы обязаны Г. Менделю. Не зная ничего ни о мутациях, ни о хромосомах, он выращивал в монастырском саду в Брюнне (Брно) садовый горошек, культивируя различные сорта, скрещивая их и наблюдая потомство в 1-м, 2-м, 3-м и т. д. поколениях. Фактически он экспериментировал с мутантами, найдя их уже готовыми в природе, и опубликовал свои результаты в 1866 г. Работа Менделя была совершенно забыта. Ее снова открыли только в 1900 г. одновременно и независимо друг от друга Корренс де Фриз и Чермак. Так это открытие стало основой совершенно новой науки — генетики. Фактически повторяя результаты Менделя, голландец де Фриз открыл, что в потомстве даже совершенно чистосортных линий появляется очень небольшое число особей (две-три на десятки тысяч) с небольшими, но скачкообразными (или дискретными — т. е. без промежуточных форм) изменениями. Он назвал это мутацией. В физике аналогичное описание механизмов скачкообразного изменения характеристик объекта свойственно квантовой теории — там тоже не наблюдается промежуточных ступеней, например, между двумя соседними энергетическими уровнями атома. Таким образом, как считал, например, Э. Шредингер, мутационную теорию де Фриза можно назвать квантовой теорией биологии.
Позже действительно было установлено, что своим происхождением мутации действительно обязаны «квантовым скачкам» в генной молекуле.
С позиций современной генетики считается, что мутации должны быть редким событием, какими они в действительности и оказываются. Вероятность возникновения вредной мутации выше, так как в природе ей соответствует больше вариантов (как и в случае вытягивания выигрышного билета в лотерее). Если бы мутации были частыми, то существенное преобладание вредных мутаций над полезными способствовало бы тому, что вместо того, чтобы улучшаться путем отбора, виды оставались бы не только неулучшенными, но, скорее всего, погибали.
Для реальной природы характерен сравнительный консерватизм биологических объектов, являющийся результатом высокой устойчивости. Природа «не любит» мутаций.
Но насколько доказательно такое постоянство биологических структур? Ответ на этот вопрос очевиден без какого-либо специального исследования. Например, почему человек, столько лет проживший в условиях холода до сих пор зимой вынужден одеваться в теплую одежду? В жизни мы также часто говорим о наследственности. Мы признаем почти абсолютным постоянство, связанное с передачей от родителя к ребенку той или иной особенность:
формы носа, длины пальцев, предрасположенности к ревматизму и т.д. В действительности из поколения в поколение, без заметного изменения в течение столетий передается весь фенотип, вся видимая и явная природа индивидуума. При этом в каждом поколении передача осуществляется материальной структурой ядер тех двух клеток, которые соединяются при оплодотворении.
Вместе с тем если мутации возникли, они могут быть унаследованы так же хорошо, как и первоначальные неизмененные признаки. Эти передаваемые потомкам мутации формально могут служить материалом для естественного отбора, как это описано Дарвином, элиминируя неприспособленных и сохраняя наиболее приспособленных.
Такие изменения ряд биологов склонны не совсем корректно называть эволюцией (микроэволюцией). Современное понятие микроэволюции — ключевое понятие эволюционных воззрений, так как оно описывает процессы, как бы реально наблюдаемые в природе. А именно считается, что микроэволюция — это распространение в популяции малых изменений в частотах аллелей на протяжении нескольких поколений, эволюционные изменения на внутривидовом уровне, которые происходят из-за мутаций, естественного отбора, искусственного отбора, переноса и дрейфа генов. Эти изменения приводят к дивергенции популяций внутри вида и, в конечном итоге, к видообразованию.
На самом же деле эти малые изменения в популяциях приводит только к образованию новых таксонов и эндемов (новых сортов пшеницы, пород коров и т. п.), но все они происходят совсем по другим причинам, а не по тем, что перечислены выше. Ни мутации, ни перенос, ни дрейф генов не имеют к этому отношения.
Очень важно отметить тот факт, что в генетический механизм наследования включен принципиально важный его элемент, существенно повышающий устойчивость всей биологической жизни, ее помехозащищенность от случайных мутаций в генах, — «консервирование» специальных половых клеток и механизм двуполого размножения, который приводит к уменьшению риска возникновения болезней и уродств.
При небольших мутациях в геномах именно половой процесс и приводит к тому, что при процессах митоза и мейоза испорченные гены и нуклеотиды родителей не попадают в новый формируемый геном ребенка, ни в соматические клетки, ни в половые, так как они расположены в разных локусах. Все возникающие у особи случайные мутации как бы автоматически проверяются на совместимость с генофондом своей популяции, а фактически с генофондом вида, и отсекаются.
Конечно, возможно возникновение мутаций и в этих половых клетках. Но чаще всего эти небольшие случайные мутации не проявляются в детях и детенышах, так как существующий механизм митоза и мейоза при делении клеток предусматривает отбраковывание испорченного нуклеотида того родителя, у которого он испорчен, хотя признаки мутантного индивидуума могут соответствовать как нормальной, так и мутантной версии. Версия, которой следует особь, называется доминантной, а не проявившаяся — рецессивной.
Аналогично мутация, которая проявляет сразу свой эффект на потомстве, называется доминантной, не проявляющая его —рецессивной.
Рецессивные мутации более часты, чем доминантные, и бывают весьма важными, хотя и не сразу обнаруживаются. Чтобы изменить свойства организма, они должны присутствовать в обеих хромосомах. Такие индивидуумы могут быть получены, когда два одинаковых рецессивных мутанта скрещиваются между собой или когда мутант скрещивается сам с собой. У гермафродитных (т. е. самоопыляющихся) растений последнее может происходить самопроизвольно. У остальных биообъектов это происходит при родственном скрещивании. Поэтому родственное скрещивание вредно.
В принципе большое количество неблагоприятных рецессивных мутаций может накапливаться и не причинять непосредственного вреда. Но они могут передаться половине потомства. Если происходит скрещивание мутантной особи с немутантной, мутацию будет нести все первое потомство, половина следующего и четверть последующего потомства, если все они вступят в брак с немутантными партнерами. Если оба партнера несут в себе более чем один скрытый недостаток, то вероятность опасности резко возрастает. Наибольшую опасность представляет скрещивание, когда мутации у партнеров совпадают. Это наиболее вероятно в первом потомстве при родственном скрещивании.
Эксперименты с растениями и животными указывают, что при родственном скрещивании, кроме сравнительно редких серьезных дефектов, имеется масса мелких, случайные комбинации которых ухудшают в целом потомство. В результате естественный отбор таких особей, даже наиболее приспособленных, ограничен и, более того, обращен в свою противоположность.
Ряд психоаналитиков, опираясь на достижения современной генетики, и, в определенной мере, следуя идеям Фрейда, считает, что при браках между родственниками, благодаря многократному наслоению одинаковых генетических характеристик, рождаются дети, у которых одни характеристики переразвиты, но за этот счет недоразвиты другие. Наиболее часто дети от таких браков страдают нервными, душевными или психическими заболеваниями. Возможно, поэтому православная церковь запрещала браки между родственниками вплоть до 7-й степени родства. Родственные браки представляют собой как бы отмирание целой социальной группы, класса или нации (как это происходит с малыми народностями, живущими достаточно изолированно от других народностей). Наилучшим лекарством против дегенерации является примесь свежей крови, когда различные классы или соседние народы и нации женятся и мешают свою кровь.
Рассматривая вопросы изменений и трансформаций видов, Дарвин допустил методическую ошибку, придав именно отбору решающее значение. Несомненно, отбор играет и в природе, и вообще в жизни большую роль, но не решающую. Отбор может только отбирать из множества существующего. Сам отбор не может выступать конструктором предметов и, как правило, не является созидательной силой. Суть естественного отбора у многих современных ученых-эволюционистов искажена: каждый видит, что в борьбе за самку побеждает самый сильный, типовой представитель этого вида в стремлении продолжить род и оставить не у дел уродливых и слабых, которые портят генофонд популяции. И сам Дарвин это иногда подчеркивал, считая, что процесс отбора в природе способствует скорее не изменению вида, а его сохранению, отбраковывая слабых и больных. Когда сильный самец побеждает слабого, больного или уродливого (из-за мутационных изменений) и продолжает род, — это способствует сохранению и породы и рода, препятствует накоплению отрицательного генетического груза в популяции, сохраняя и предохраняя генотип от всяких серьезных изменений и тем препятствует всякой биологической эволюции. Кишечная палочка так и остается кишечной палочкой, дрожжи — дрожжами, а лягушки — лягушками.
С позиций генетики наследственность — явление полностью противоположно понятию биологической эволюции. Наследственность — это передача признаков родителей или родительских признаков детям, а биологическая эволюция — процесс появления новых видов от существующих, за счет приобретения детьми признаков, которых не было у родителей. Считается, что в мире нет двух одинаковых особей. Но различия и отличия детей от родителей есть обязательное проявление законов наследственности (физической эволюции), и все признаки, которые есть у детей, они получили от родителей в определенных сочетаниях.
На самом деле в природе до сих пор не зафиксирована ни одна так называемая положительная мутация. Единственным примером такой мутации может служить ограниченное число образования подвидов при явлении полиплоидии. Но и в этом случае новый вид не образуется. Механизм наследственности свидетельствует не о предрасположенности биологического мира к эволюционным изменениям, а о его консервативности и борьбе с любыми изменениями, противодействию им и защите от них. Именно на эти противодействия нацелена и еще одна важнейшая система животного и растительного мира — иммунная система каждого организма. Чтобы допустить серьезные эволюционные изменения в исходном организме, она однозначно должна быть сильно ослаблена или отсутствовать вообще (например, при заболевании СПИДом). Но если даже и происходят существенные мутации в исходных геномах, то они приводят не к появлению нового вида, а к рождению мутантов и уродов, детей с патологиями и болезнями, к бесплодию и т. д.
В современной эволюционной биологии вопросы видообразования до сих пор остаются не решенными. Все известные виды появлялись как бы внезапно, т. е. через бифуркацию. До сих пор в раскопках (палеонтологических летописях) не обнаружено ни одного «промежуточного» вида, свидетельствующего о постепенном, плавном (континуальном) появлении новых видов в процессе физической эволюции. Все видовые ряды являются конечными и дискретными. Таким образом, реально наблюдается типично квантовая природа смены видов.
3. Генная инженерия - перспективы и последствия.
Генная инженерия — совокупность приемов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие организмы.
Генная инженерия — под этим термином в средствах массовой информации обычно подразумевают методы искусственного воздействия на гены, с целью какого-либо видоизменен
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|