- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Подтверждения
Использование питательных веществ и реминерализация
Уолтер К. Доддс , Мэтт Р. Уайлс , « Пресноводная экология» (второе издание) , 2010
Парадокс ограничения планктона и питательных веществ
Парадокс в планктоне . Хотя его можно рассматривать как «соломенного человека», учитывая то, что в настоящее время известно о водной экологии , предлагаемый парадокс является полезной отправной точкой для обсуждения и, возможно, служит основой для наиболее распространенного вопроса, который задавался в выпускных экзаменах по водной экологии в прошлом. 40 лет
принцип конкурентного исключения ) ксообществам фитопланктона . Конкурентное исключение происходит потому, что только один вид может быть лучшим конкурентом для одного ограничивающего ресурса. В среде, в которой несколько видов конкурируют за один ресурс, превосходящий конкурент в конечном итоге истребит другие.
Хатчинсон утверждал, что, поскольку озера представляют собой очень хорошо перемешанную среду, ограничивающие питательные вещества хорошо смешиваются и в равной степени доступны для фитопланктона. Учитывая, что большинство клеток имеют сходные требования и конкурируют за одни и те же питательные вещества, принцип конкурентного исключения должен ограничивать количество видов, присутствующих в любое время. Парадокс, который он отметил, заключается в том, что типичное мезотрофное или олиготрофное озеро содержит много видов (обычно 10–100) фитопланктона, которые присутствуют одновременно.
Объяснения разнообразия планктона включают следующее: (1) хищничество зоопланктоном и вирусами удаляет или подавляет доминирующих конкурентов ( Suttle et al. , 199 Харрис, 1980), (2) импульсы и микропатчи питательных веществ от неравномерного смешивания и выделения приводят к неравновесным условиям (обсуждается далее в этой главе), (3) взаимные или полезные взаимодействия способствуют другим конкурентам, уступающим, (4) многие озера не находятся в равновесных условиях в течение периода времени, превышающего 1 месяц, и время, необходимое для доминирующих видов фитопланктона, чтобы победить конкурентов, составляет более одного месяца ( ). Идея различных конкурентных способностей привела к теории соотношения ресурсов и определению того, как соотношение Redfield связано с ограничением питательных веществ.6), (5) разные конкурентные способности приводят к тому, что разные питательные вещества ограничивают разные виды, и (6) хаос (в математическом смысле) возникает, когда виды конкурируют за три или более ресурсов ( Huisman and Weissing, 1999
Просмотр главыКнига покупок
Использование питательных веществ и реминерализация
Уолтер К. Доддс , Мэтт Р. Уайлс , « Пресноводная экология» (третье издание) , 2020 г.
Парадокс ограничения планктона и питательных веществ
Парадокс в планктоне . Хотя мы можем рассматривать парадокс как «соломенного человека», учитывая то, что в настоящее время известно об экологии водных ресурсов , предлагаемый парадокс представляет собой полезную отправную точку для обсуждения и, возможно, служит основой для наиболее распространенного вопроса, задаваемого на выпускных экзаменах по экологии водных ресурсов в последние 50 лет
принцип конкурентного исключения ) ксообществам фитопланктона . Конкурентное исключение происходит потому, что только один вид может быть лучшим конкурентом для одного ограничивающего ресурса. В среде, в которой несколько видов конкурируют за один ресурс, превосходящий конкурент в конечном итоге истребит другие. Применение ЛейбигЗакон и принцип конкурентного исключения требуют нескольких допущений: (1) все питательные вещества хорошо перемешаны во времени и пространстве в окружающей среде (равновесие), (2) все организмы имеют примерно одинаковые требования, и (3) конкуренция имеет время, чтобы вызвать ухудшение конкуренты исчезают из сообщества. Хатчинсон утверждал, что, поскольку озера представляют собой очень хорошо смешанную среду, большинство аспектов этих допущений должны соблюдаться. Парадокс, который он отметил, заключается в том, что типичное мезотрофное или олиготрофное озеро содержит много видов (обычно 10–100) фитопланктона, которые присутствуют одновременно.
Существует множество объяснений высокого планктонного разнообразия, предложенного Хатчинсоном и другими. (1) Хищничество зоопланктоном и вирусами устраняет или подавляет доминирующих конкурентов ( Suttle et al., 1990 Harris, 1986). (2) Импульсы и микропатчи питательных веществ из-за неравномерного смешивания и выведения приводят к неравновесным условиям (обсуждается далее в этой главе). (3) Мутуалистические или полезные взаимодействия способствуют другим конкурентам. (4) Многие озера не находятся в равновесных условиях в течение периода времени, превышающего месяц, и время, необходимое для доминирующих видов фитопланктона, чтобы победить конкурентов, составляет более месяца ( ). Идея различных конкурентных способностей привела к теории соотношения ресурсов и определению того, как соотношение Redfield соотносится с ограничением питательных веществ.). (5) Различные конкурентные способности приводят к тому, что разные питательные вещества ограничивают разные виды. И (6) хаос (в математическом смысле) возникает, когда виды конкурируют за три или более ресурсов ( Huisman and Weissing, 1999
Просмотр главыКнига покупок
Конкурс ресурсов и экологический успех фитопланктона
ЕЛЕНА LITCHMAN в эволюции первичных продуцентов в море , 2007
VI ВЫЗОВЫ И БУДУЩИЕ НАПРАВЛЕНИЯ
Экология фитопланктона, которая достигла значительных успехов за последние несколько десятилетий и поставила, а также ответила на многие фундаментальные вопросы экологии, такие как проблема конкурентного исключения и сосуществования видов, сформулированная в парадоксе Хатчинсона о планктоне (
Динамическое регулирование использования ресурсов и конкурентоспособности
Модели конкуренции ресурсов в фитопланктоне обычно предполагают постоянные значения параметров для каждого вида (например, Grover 1991c ; Huisman and Weissing 1994 ; Ducobu et al. 1998) . Тем не менее, многочисленные экспериментальныеИсследования показывают, что различные параметры, которые определяют конкурентную способность, такие как зависящие от питательных веществ параметры поглощения и роста (например, см. уравнение 3 V max , может изменяться в несколько раз в зависимости от степени ограничения питательности или интенсивности облучения (см. Harrison et al. 1989) ; минимальная квота питательных веществ, Q min , для азота также зависит от уровня освещенности ( Rhee and Gotham 1981) зависят от условий роста и, следовательно, представляют собой не просто число, а распределение вокруг среднего значения. Например, максимальная скорость потребления питательных веществ , . Однако существуют случаи, когда простые модели кажутся неадекватными, флуктуирующие условия могут вызвать физиологические реакции, которые являются достаточно значительными, чтобы вызвать разногласия между моделями и экспериментами (например, ). Механизмы динамической регуляции сбора и использования ресурсов в фитопланктоне начинают изучаться с помощью геномного и метаболического подходов ( . Понимание динамики акклиматизации). Часто эти зависимости, не включенные в модель, не влияют на динамику, и исход соревнования и модели с постоянными коэффициентами достаточны для прогнозирования конкурентных взаимодействий ( Tilman 1977 ; Huisman et al. 1999a) Grover 1991a , b , c , 1992 ; Litchman 2003 Armbrust et al. 2004 ; Allen 2005 ; Parker and Armbrust 2005 ). Повышение или понижение регулятора потребления питательных веществ и механизмов фотосинтеза происходит быстро при различной степени ограничения ресурсов (например, Hildebrand и Dahlin 2000 ; Song and Ward 2004 ; Allen et al. 2005 ; Hildebrand 2005 ) и может изменить конкурентоспособность видов способность ( Klausmeier et al. 2007) фитопланктона для изменения полей ресурсов и определения того, как такая акклиматизация может повлиять на конкурентоспособность ресурсов и результаты конкуренции, представляет собой новую проблему и улучшит нашу способность прогнозировать динамику фитопланктона в целом.
B Ресурс Взаимодействие
Большинство моделей конкурентной борьбы за ресурсы часто предполагают отсутствие взаимодействия между ресурсами и описывают скорость роста фитопланктона как минимальную функцию от всех потенциально ограничивающих ресурсов (закон минимума Либиха или, точнее, упоминается как ограничение Блэкмана, если рассматриваются ставки, см. Cullen 1991). Wheeler et al. 1983 ; Kudela and Cochlan 2000). Параметры роста и поглощения для каждого ресурса предполагаются независимыми от других ресурсов в моделях. На самом деле физиология соответствующих процессов является более сложной. Экологи фитопланктона давно знают, что поглощение питательного вещества зависит от света и других питательных веществ, часто специфическим для вида образом (например,). Поглощение нитрата ингибируется аммонием, и такое ингибирование может значительно изменить использование нитрата ( Syrett 1981 ; Dortch 1990 ). Скорость поглощения нитратов, фосфатов и других питательных веществ также зависит от интенсивности облучения, и, поскольку эта зависимость зависит от вида, изменения в световом режиме могут способствовать конкуренции питательных веществ ( Ahn et al. 2002 ; Litchman et al. 2004), Нужно ли учитывать такие отношения между ресурсами, чтобы адекватно моделировать конкурентные взаимодействия? Модель, которая включала взаимодействие света-аммония-нитрата, использовалась для успешного прогнозирования динамики нитратов в океане с высоким содержанием питательных веществ и низким содержанием хлорофилла (HNLC), что предполагает, что такие модели могут быть более эффективными в случаях колимитации ресурсов ( Armstrong 1999 ).
С Эволюция конкурентоспособности
Наряду с кратковременной гибкостью и пластичностью способности фитопланктона приобретать и конкурировать за ресурсы, конкурентная способность фитопланктона, вероятно, развивается в ответ на изменения в окружающей среде как благодаря биотическим, так и абиотическим селективным силам. Из-за большого числа популяций, короткого времени генерации и клонального размножения фитопланктон может эволюционировать в относительно быстрых временных масштабах ( Lynch et al. 1991 ; Yoshida et al. 2003 , 2004 ). Мутанты или клоны с лучшей конкурентоспособностью благодаря изменению экофизиологических параметров будут отбираться при данных условиях окружающей среды.Однако из-за компромиссов между различными параметрами, а также между конкурентными способностями для различных ресурсов и других компонентов пригодности, таких как устойчивость к травлению, эволюция, вероятно, будет ограничена, так что появление супервидов маловероятно. Быстрое развитие конкурентоспособности может изменить динамику и результаты конкуренции за ресурсы и взаимодействия в пищевой сети ( ). Очень мало известно о том, как быстро может развиваться конкурентоспособность и различаются ли темпы эволюционного изменения конкурентоспособности среди таксономических групп или для разных ресурсов. Лабораторные эксперименты с зеленой водорослью Chlamydomonas Chlorella zooplankton, пасущегося под давлением привели к отбору значительно более устойчивых к травлению клонов с низкой конкурентоспособностью питательных веществ (
Существующие модели конкурентной борьбы за ресурсы в фитопланктоне не учитывают возможную эволюцию конкурентной способности и появление новых фенотипов и, следовательно, могут не быть в состоянии реально описать динамику конкурентных сообществ. Перспективным подходом к моделированию эволюции в экологическом контексте, который учитывает сложные взаимодействия видов, является подход адаптивной динамики, где приспособленность является функцией частот взаимодействующих генотипов ( Geritz et al. 1998 ; Waxman and Gavrilets 2005). Этот подход расширяет методы эволюционной теории игр и может использоваться для изучения того, какие новые конкурентные стратегии и эволюционные линии могут появиться в различных условиях окружающей среды (Доебели и Дикманн 2005 ; Кисди и Джилленберг 2005 ; Ваксман и Гаврилец 2005 ).
D Филогенетические отношения
Экспериментальные данные указывают на то, что фитопланктон из основных таксономических групп отличается по своей конкурентной способности к различным ресурсам (питательным веществам и свету). Различия в конкурентных способностях и стратегиях между таксономическими группами могут частично объясняться условиями окружающей среды в момент их возникновения ( Фальковский и др. 2004 ; Катц и др. 2004 ; Тоцци и др. 2004 ; Личман и др. Song and Ward 2004 ; LitchmanОтправлено). В то же время молекулярные и физиологические данные указывают на то, что в основных таксонах виды морских и пресноводных сред отличаются в своих стратегиях использования питательных веществ ( представлено). Например, транскрипция азотредуктазы в морских зеленых водорослях регулируется по-разному по сравнению с пресноводными зелеными водорослями, что может быть связано с различиями в степени ограничения нитратами в морских и пресноводных системах ( . Понимание влияния филогенетических связей в сравнении с экологическими факторами на фитопланктон Конкурентные способности ставят новые интересные задачи, которые требуют синергизма генетического, физиологического и экологического подходов.и другие. Song and Ward 2004 ). Даже генетически близкие клоны, обитающие на разных глубинах, могут различаться по своей питательной или световой физиологии и конкурентным способностям ( Moore et al . 1998 ; Rocap et al. 2003 ; Stomp et al. 2004)
E Заключительные замечания
Конкурс ресурсов - это фундаментальный экологический процесс, который структурирует сообщества фитопланктона. За последние несколько десятилетий были достигнуты значительные теоретические и экспериментальные успехи в области конкуренции за ресурсы фитопланктона. Тем не менее, мы только начинаем понимать, как различия в конкурентных стратегиях между основными функциональными группами, а также отдельными видами, способствуют разнообразию фитопланктона и изменению сообщества в природе. Разнообразные аспекты конкуренции за ресурсы, такие как механистические компромиссы в конкурентных способностях, а также их гибкость и эволюция, а также конкуренция в условиях переменнойнеобходимо учитывать условия, если мы хотим понять и / или предсказать структуру и функцию прошлого, настоящего и будущего сообществ фитопланктона.
Подтверждения
Работа над этой главой была частично поддержана грантами NSF OCE-0084032, DEB0445265 (0610531) и DEB-0610532. Комментарии CA Klausmeier, PG Falkowski и AH Knoll приветствуются. Это вклад Биологической станции Келлогг. 1424.
Просмотр главыКнига покупок
Микробная Экология
Н. Д. Грей , И. М. Руководитель , в Энциклопедии экологии , 2008
Связь микробного разнообразия с экологическими механизмами и факторами окружающей среды
структура микробного сообщества экологов в отношении мелкомасштабных градиентов света, температуры, солености , акцепторов электронов и доноров, а также связь генетически различимых организмов с конкретными условиями окружающей среды. Эти исследования непосредственно затрагивают такие вопросы, как « парадокс планктона » Хатчинсона , который ставит под сомнение аномальное сосуществование видов, якобы использующих одни и те же ресурсы в явно однородных средах без конкурентного исключения. Сообщества прохлорофитов (кислородсодержащие фотосинтетические бактерии) в поверхностных водах океана структурированы по интенсивности света , тогда как сосуществующие гигантские сероокисляющие бактерии из рода вПресноводные отложения структурированы по редокс-градиентам (рис. 5 ) и их слегка различаются требования к нитратам.
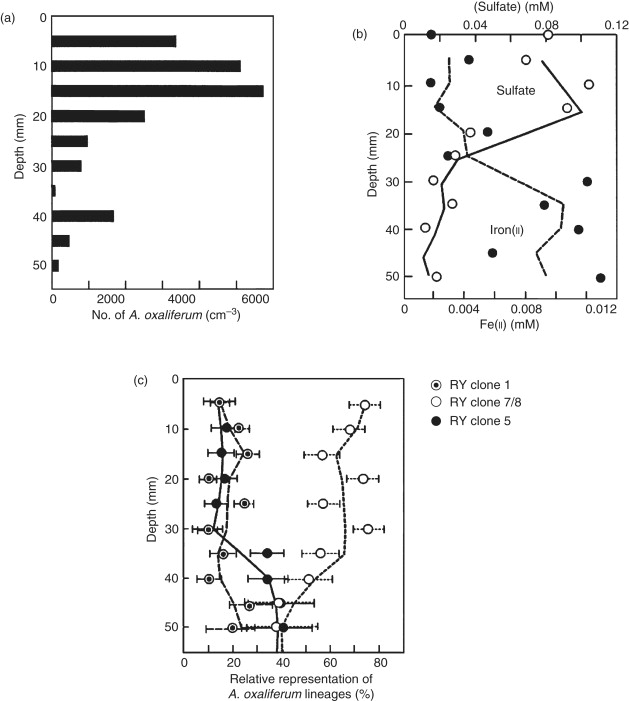
Войдите, чтобы скачать полноразмерное изображение
На рисунке 5 . (a) Профили глубины клеток Achromatium , (b) окислительно-восстановительные химические вещества (Fe 2+ и SO 4 2- ) и (c) относительная численность трех линий Achromatium, идентифицированных с использованием FISH в ядре осадка от Rydal Water, ВЕЛИКОБРИТАНИЯ. Линии тренда показывают трехточечную скользящую среднюю данных, а столбцы ошибок представляют 95% доверительные интервалы для подсчетов субпопуляций Achromatium . Воспроизводится из Grey ND, Howarth R, Rowan A, et al. (1999). Природные сообщества Achromatium oxaliferum включают генетически, морфологически и экологически различающиеся субпопуляции .Прикладная и экологическая микробиология 65: 5089–5099.
Хотя некоторые факторы окружающей среды или возмущения играют роль в поддержании разнообразия, например, разнообразие растений и рост растений в почве, другие вызывают потерю разнообразия, например, управление земельными ресурсами и загрязнение окружающей среды. Высокая степень экологической избыточности, по-видимому, является характерной чертой микробных сообществ ; следовательно, потеря разнообразия сама по себе не обязательно влияет на функционирование экосистемы, но разнообразие и функциональная избыточность, по-видимому, важны для поддержания устойчивости и устойчивости к экологическим нарушениям.
Неоднородность среды обитания издавна использовалась для объяснения более высокого разнообразия видов микробов в неоднородных средах, таких как почвы. Сравнение показателей видового богатства и разнообразия на основе анализа последовательностей 16S рРНК, полученных из проб окружающей среды, показало, что пространственная изоляция и разнородность ресурсов играют важную роль в поддержании бактериального разнообразия. В почвах с низким содержанием влаги существуют бактериальные сообщества с более высоким разнообразием, чем в почвах с высоким содержанием влаги, где существует большая связь и возможности для конкуренции между сосуществующими бактериями.
Пространственные закономерности в распределении видов долгое время были основой общей экологии, Тем не менее, в микробном мире эта область полна противоречий. Это главным образом является результатом предположения о том, что малый размер клеток, большой размер популяции и простота расселения приведут к тому, что микробные таксоны будут иметь космополитическое распределение. Соответственно, структура местного микробного сообщества является результатом выбора конкретных таксонов на основе экологической пригодности из глобального пула микробных видов. Эта гипотеза была впервые предложена в 1934 году Бассом Бекингом и подытожена фразой «все везде, окружающая среда выбирает». Отсюда следует, что функциональное разнообразие также должно иметь повсеместное распространение. Эта точка зрения подтверждается наблюдением, что большая доля мирового разнообразия инфузорий и других простейшихможно обнаружить при любом обширном обследовании местного разнообразия. Эти исследования были основаны на выявлении морфоспецифических видов, которые, как известно, скрывают богатство генетического разнообразия . Существуют убедительные доказательства появления эндемичных прокариот в почве, горячих источниках и других средах. Аргументы за и против возникновения эндемических микроорганизмов омрачены классическими моделями аллопатрического видообразования, основанными на географической изоляции.из двух популяций с последующим генетическим расхождением и в конечном итоге видообразованием. На самом деле это частный случай более общей формулировки, где аллопатрическое видообразование может произойти, если скорость генетической дивергенции больше скорости рассеивания. Эта общая модель может объяснить возникновение эндемизма у микробных видов даже при отсутствии явных географических барьеров для распространения. Вероятно, что, как и в случае с другими организмами, будут глобально распространенные организмы, которые являются более или менее космополитическими и более редкими организмами, которые с большей вероятностью будут демонстрировать более быструю эволюцию, чем рассеяние, и, следовательно, проявлять эндемизм.
Просмотр главыКнига покупок
Пресноводные экосистемы
Уолтер К. Доддс , Мэтт Р. Уайлс , « Пресноводная экология» (второе издание) , 2010
Биоразнообразие и функция экосистемы
Биоразнообразие экосистем может быть связано с темпами экосистемных процессов и биологической структурой экосистем различными способами. Например, из-за разнообразия видов (разнообразия видов сетчатых летающих насекомых фильтруют более высокие порции взвешенных частиц из толщи воды, чем монокультуры) ( обнаружены отрицательные связи между разнообразием видов и разложением опавших листьев, поскольку Cardinale et al. , 2002 ). Однако отношения между биоразнообразием и функциями экосистемы могут быть переменными и сложными. Изучение функционально и конкурентно доминирующих листопадных Pycnopsyche опавших Pycnopsycheуменьшает присутствие других видов измельчителей (сильный конкурент) и разлагает большое количество мусора и, таким образом, является функционально доминирующим ( Creed et al. , 2009 ).
Учитывая нынешний кризис биоразнообразия, особенно в пресноводных средах обитания, усилия по установлению взаимосвязей между биоразнообразием и функционированием экосистем функционально избыточнымибыли центральной проблемой в пресноводной экологии (Шульце и Муни, 1994). Первая проблема заключается в том, как определить функцию экосистемы. Функция может относиться к показателям основных процессов, таких как фотосинтез, дыхание, денитрификация или задержка фосфора. Это может также относиться к более конкретным вещам, таким как производство растительной биомассы для травоядных. Суть этого вопроса заключается в том, если видыв отношении их роли в экосистеме (т. е. могут ли виды быть удалены из сообщества без изменения в конкретном экосистемном процессе?). Функциональная избыточность может варьироваться в зависимости от рассматриваемого экосистемного процесса, конкретной среды обитания и времени. У многих континентальных сред обитания есть разнообразные сообщества со значительной избыточностью.
Связь между разнообразием и экосистемными процессами, приводящими к круговороту питательных веществ, неясна, потому что мало что известно о микробном разнообразии и степени избыточности конкретных функциональных групп ( Meyer, 1994). Хотя Хатчинсон привел ранние аргументы о функциональной избыточности фитопланктона парадокс планктон (см.
Разнообразие бентических животных может оказывать сильное влияние на процессы обмена веществ между толщей воды и бентической зоной ( ), и различные виды по-разному изменяют поток питательных веществ (например, рытье туннелей, рытье туннелей, разжигать отложения, активно закачивать насыщенную кислородом воду в отложения) и обработка различных типов донных материалов). Исследование тропической реки с разнообразным скоплением рыб, обсуждавшееся ранее, показало, что потеря только одного вида рыб, крупного мигрирующего детривора , значительно снизила перенос углерода вниз по течению ( толщей Covich et al. , 1999 ). Бентические животные могут играть важную роль в потоках энергии и круговороте питательных веществ ( Рис. 24.5 Taylor et al. , 2006). Это гениальное исследование использовало забор вдоль середины ручья и исключало или включало интересующую рыбу. Данные свидетельствуют о том, что признаки одного вида могут изменять функцию экосистемы даже в разнообразной системе, где функциональная избыточность, как ожидается, будет высокой.
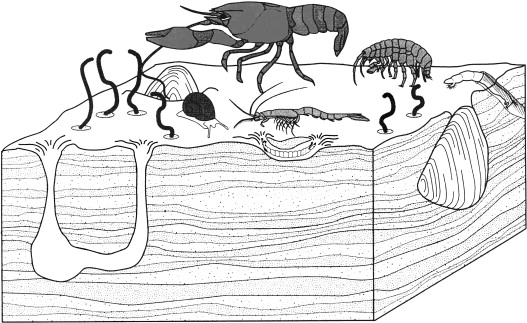
Войдите, чтобы скачать полноразмерное изображение
Рисунок 24.5 . Бентические макробеспозвоночные, которые роются в слоистых отложениях и ускоряют круговорот питательных веществ и движение материалов в толщу воды. При закапывании двустворчатых моллюсков, раков, трубчатых червей и личинок водных насекомых O 2 смешивается с осадками в результате своей деятельности. Поверхностные беспозвоночные увеличивают оборот микробных сообществ и увеличивают осадочную взвесь.
(Воспроизведено с разрешения Covich et al. , 1999 ).
Некоторые из лучших данных для функционального разделения экосистемных процессов, связанных с биоразнообразием, получены в результате исследований двух видов креветок, которые разлагают опавшие листья в ручьях Пуэрто-Рико ( но при этом они очищают микробы от листьев и более эффективно фильтруют мелкие частицы из толщи воды. Транспортировка твердых частиц наиболее высока в ручьях, в которых оба вида креветок редки из-за присутствия хищных рыб ( ). Пример Пуэрто-Рико является лишь одним из примеров того, как содействие среди различных видов может опосредовать воздействие видов на экосистемные процессы, и этот вывод был воспроизведен в болотных растениях ( опавшие Covich, 1997 ). Оба вида ( Xiphocaris elongata и Atya lanipes ) могут разлагать опавшие листья, но их разрушение значительно более эффективно, и потоки более задерживают органические частицы, когда присутствуют оба вида. Атя не ломает целые листья так быстро, как Xiphocaris Pringle et al. , 1999 ). Такая взаимосвязь между биоразнообразием и функционированием экосистем может встречаться и в материковых потоках ( Jonsson et al. , 2001 Bertness and Hacker, 1994 ; Bertness and Leonard, 1997 ).
Различия в структуре или функции экосистемы также могут быть связаны с разнообразием ( Франция и Даффи, 2006 Morin and McGrady-Steed, 2004 ).). Эксперименты с множеством микробных сообществ, собранных по-разному, позволяют предположить, что изменение частоты дыхания связано с биоразнообразием. Более разнообразные сообщества были более схожими по частоте дыхания, но менее разнообразные сообщества различались более широко (
Поскольку многочисленные исследования были опубликованы, стал возможен более глубокий метаанализ взаимосвязей между разнообразием и функциями экосистемы. Отчет по заказу Экологического общества Америки ( Hooper et al. , 2005) выразил уверенность в том, что (1) функциональные характеристики видов влияют на экосистемы, (2) исчезновение и интродукция видов, вызванных человеком, оказали влияние на экосистемы способами, которые являются дорогостоящими для человека, (3) последствия потери видов или изменения структуры сообщества на экосистемах в зависимости от местных условий, (4) некоторые свойства экосистемы не связаны с разнообразием из-за функциональной избыточности, и (5) по мере увеличения пространственной и временной изменчивости требуется больше видов для создания стабильных поставок экосистемных товаров и услуг. Анализ 446 наземных и водных исследований выявил наиболее сильное влияние на численность первичного производителя и первичного потребителя, а также на активность разложения ( Balvanera et al. , 2006). Анализ 111 исследований показал, что идентичность видов является единственным наиболее важным детерминантом воздействия экосистем на потерю видов ( Cardinale et al. , 2006 ).
Просмотр главыКнига покупок
Взаимодействие бентических водорослей с их субстратами
Джоанн М. Буркхолдер , « Экология водорослей» , 1996 г.
V. ВЫВОДЫ И РЕКОМЕНДАЦИИ
Сожительства одноклеточных или многоклеточных организмов различного систематического положения реализуется в виде непрерывной серии ассоциаций, из сыпучих nonobligate ectosymbioses к высокоразвитых облигатных endosymbioses ( ). Этот спектр взаимодействий встречается среди донных водорослей и их колонизированных субстратов. Хотя многие виды водорослей являются универсалами среды обитания или случайными колонизаторами различных живых и неживых поверхностей ( ), некоторые демонстрируют более специализированные субстратные предпочтения при необлигатных эктосимбиозах; и другие становятся тесно биохимически связанными «партнерами» в мутуалистических эндосимбиозах с растениями-хозяевами , грибами или животными ( Kies, 1992 Whitton, 1975 ; Round, 1981 Meeks et al., 1985). Многие из этих взаимодействий, особенно при рыхлых эктосимбиозах, по-видимому, включают небольшие концентрации плохо охарактеризованных веществ, выделяющихся из субстратов.
Несмотря на значительные исследования в области экологии донных водорослей , наши знания о спектре химических веществ, управлении биохимическими сигналами и общей значимости взаимодействий донных водорослей / субстрата с пресноводными экосистемами остаются ограниченными, в основном из-за ограничений в доступной технологии микромасштаба . Как показало применение кислородных микроэлектродов, микроавторадиографии, молекулярных зондов и других методологий за последнее десятилетие, реалистичное рассмотрение химических взаимодействий с участием донных водорослей должно все больше фокусироваться на масштабах возникновения, а именно на химических микроградиентах среды обитания микроничей.
Микромасштабные методы и подходы постоянно позволяют разрешать очевидные парадоксы . знаменитый « парадокс планктона » - классическая иллюстрация этого момента; парадокс многих видов водорослей в «одной» видимой среде обитания был разрешен путем признания соответствующих микропространственных и временных масштабов планктона ). Микромасштабные измерения скорости тока позволили более точно описать физические / химические эффекты пограничного слоя на сообщества донных водорослей и химическую микросреду в матрице перифитонной биопленки ( C или кислородный дисбаланс Стивенсон, 1983 ; Рейтер и Карлсон, 1986 ; Стивенсон и Гловер, 1993 ) , Микроэлектроды позволили реалистично оценить продуктивность перифитона и избежать диффузионного 14 Carlton and Wetzel, 1987 ; см. Раздел IIIпроблемы, встречающиеся с более традиционными методами ( ). Микроэлектроды, используемые с молекулярными методами, также позволили обнаружить такие процессы, как фиксация азота на микробитатах частиц детрита , информация, которая перенаправила парадигмы об общем или макромасштабном функционировании вод с ограниченным содержанием азота ( ). Microautoradiography позволило сосредоточиться на химической микросреде , имеющей отношение к эпифитным микроводорослям и переворачивается представлениями исследователей о важности незначительных потерь P от макрофитов ткани как значительные ( ), питательных веществ для эпифитов . Определение механизмов распознавания клеточной поверхности среди хозяев и Paerl, 1990 ; Currin et al., 1990 Moeller et al., 1988 ), а не пренебрежимо мало ( Moeller et al., 1985).Эндосимбионты были за пределами нашей досягаемости до появления зондов, конфокальной микроскопии и связанных с ними молекулярных приложений, которые позволили сосредоточиться на соответствующем микроскопическом масштабе, чтобы начать разрешать биохимические и молекулярные взаимодействия ( появились ДНК / РНК White et al., 1987 ; Reisser, 1992b ).
В дополнение к продолжающемуся развитию и использованию микромасштабных методов, необходимы новые концептуальные подходы для выявления взаимодействия бентических водорослей и субстрата. Существует определенное знакомое утешение в использовании устоявшихся парадигм или догм для определения гипотез и экспериментов, касающихся физиологической экологии бентических водорослей. Хотя в некоторой степени такие мыслительные процессы неизбежны, опасность в этом комфорте заключается в том, что основные контролирующие механизмы и дажеглавные игроки в этих средах обитания могут быть полностью пропущены. Предыдущие результаты часто основаны на методах, которые с течением времени становятся признанными ограниченными или неадекватными для предыдущего применения. Ирония в том, что, соблюдая то, что «в общем» известно как истинное, мы иногда не видим, что на самом деле верно для более конкретного случая или альтернативной ситуации.
Например, парадигма низкой гетеротрофной способности сообществ микроводорослей, вероятно, выходящая за рамки намерений авторов, была основана на сравнительном поглощении двух растворенных органических субстратов водорослями и бактериями в поверхностных водах олиготрофных озер ( Wright and Hobbie, 1966 Gaines and Elbrächter, 1987 ; Sanders and Porter, 1988 ; Tranvik et al., 1989 ; Carrick and Fahnensteil, 1990) и некоторых пресноводных фитопланктонах. также было обнаружено эффективное использование естественных уровней растворенной органики (). Более поздние исследования установили, что фаготрофный фитопланктон играет важную роль в «микробной петле» олиготрофно-эвтрофных поверхностных вод ( ). Если было обнаружено , что фитопланктон в более изобильных средах с низким содержанием органических субстратов использует гетеротрофию , разумно ожидать, что бентические микроводоросли также могут полагаться на него, по крайней мере факультативно (например,Гейнс и Эльбрахтер, 1987 ; Lewitus and Caron, 1991 Pfiester and Popovský, 1979 ; также см. Tuchman, Глава 10 , этот том). Значение гетеротрофии в химических взаимодействиях донных водорослей - вопрос, заслуживающий тщательной оценки; Частичные или растворенные органические ресурсы, вероятно, окажутся важными для многих донных микроводорослей в получении углерода и других питательных веществ из связанных с ними субстратов.
Наконец, концептуальные подходы к пониманию химического взаимодействия бентических водорослей с их субстратами будут и впредь совершенствоваться путем пересечения пресноводной / морской «границы», которая обычно вводится при формальной подготовке ученых-водников. Бентические водоросли встречаются вдоль континуума от озер и рек до лиманов и прибрежных морских вод, и, что неудивительно, эти сообщества имеют много общих черт ( ). Пресноводная экология, в свою очередь, внесла классические выводы о биологии некоторых основных групп донных водорослей [например, динофлагеллят ; ) которые были найдены аналогичными в морских средах обитания (например, Раунд, 1981 ; Буркхолдер и Ветцель, 1989b ; Эверитт и Буркхолдер, 1991 ). О некоторых классических исследованиях физиологической экологии донных водорослей сообщили морские системы, например, с прямыми аналогиями с пресноводными местообитаниями (например, Harlin, 1973 ; Sand-Jensen, 1977 ;Penhale and Thayer, 1980 ; Салливан и Монкрейф, 1990Pfiester and Popovský, (1979) ], приложения для микротехники и химические взаимодействия водорослей и субстрата (например, Carlton and Wetzel, 1988 ; Moeller et al., 1988 Coleman и Burkholder). 1995 ; Burkholder et al., 1995). Учитывая, что наши знания о взаимодействии водорослей и субстратов во многих отношениях отсутствуют или ограничены, наше понимание будет укрепляться и расширяться благодаря пониманию экологии донных водорослей в водных экосистемах .
Просмотр главыКнига покупок
Интегративная экология: от молекул к экосистемам
Роберт Ptacnik , ... Хельмут Хиллебранд , в достижении экологических исследований , 2010
Я Введение
Растущее осознание растущей утраты глобального биоразнообразия ( Worm et al. , 2006 ) способствовало значительному сдвигу в экологических исследованиях в последнее десятилетие или около того. Первоначально исследователи интересовались главным образом тем, как разнообразие регулируется в природных сообществах и как, по-видимому, сходные виды могут сосуществовать, но в настоящее время основное внимание уделяется пониманию воздействия разнообразия на экосистемные процессы и услуги ( Hillebrand and Matthiessen, 2009; Hooper et al ., 2005; Reiss et al ., 2009 ).
Начиная с экспериментов с семенными лугами Тильмана ( ), исследования в области взаимосвязей между биоразнообразием и функционированием экосистем (B – EF) быстро развивались, особенно в экологии суши ( ). В водной среде обитания большая часть экспериментальной работы на сегодняшний день была сосредоточена на взаимосвязях B – EF в микробных микрокосмах (например ) или, чаще, в бентической макрофауне (например ), с очень небольшим количеством исследований, включающих как микро-, так и макроорганизмы. (но посмотри Тилмана Tilman et al. 1996 Hooper et al. , 2005 Petchey et al ., 1999 Perkins et al ., 2010 Reiss et al ., 2010b ). Бентические сообществаво многих отношениях гораздо более похожи на наземные сообщества, чем их пелагиальные аналоги, которым до сих пор уделялось меньше внимания в исследованиях B – EF. Фактически, только 7 из 84 исследований в наборе данных синтеза, собранных Cardinale et al. (2006b) имеют дело с пелагическими организмами, и это все лабораторные, а не полевые эксперименты. Тем не менее, эта экспериментальная работа с искусственными сообществами планктона сыграла ключевую роль в процессе перехода от раннего внимания к луговым сообществам и основным производителям к тому, как разнообразие влияет на трофи
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|