- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
ДВИГАТЕЛЬНЫЕ СИСТЕМЫ. ОБЩИЙ ПЛАН ОРГАНИЗАЦИИ
ДВИГАТЕЛЬНЫЕ СИСТЕМЫ. ОБЩИЙ ПЛАН ОРГАНИЗАЦИИ
Каждый нейронный механизм, участвующий в регуляции фазной и позной мышечной активности, по предложению ряда физиологов, называется двигательной системой. Поэтому когда говорят: «двигательная система спинного мозга», то подразумевают все механизмы спинного мозга, участвующие в процессах регуляции мышечной активности. Аналогично, можно говорить о двигательных системах ствола мозга, подкорковых структур, мозжечка, коры больших полушарий.
В спинном мозге расположена основная структура — альфа-мотонейрон, аксон которого является единственным каналом, соединяющим нервную систему со скелетной мышцей. Только возбуждение альфа-мотонейрона приводит к активации соответствующих мышечных волокон. В спинном мозге существует два механизма, активирующих альфа-мотонейроны. 1-й механизм — это прямое нисходящее влияние на альфа-мотонейрон, например, такие воздействия могут оказывать некоторые аксоны гигантских пирамидных клеток Беца, расположенных в двигательной коре. Однако в мозге чаще активация альфа-мотонейрона осуществляется опосредованно, через дополнительные нейроны — вставочные, число которых в спинном мозге огромно. Можно также возбудить альфа-мотонейроны за счет 2-го
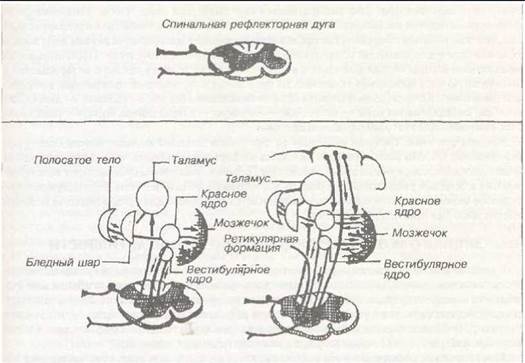
Рис. 19. Конвергенция на спинальном мотонейроне влияний от вышележащих структур» ответственных за тонус и локомоцию.
механизма активации с помощью гамма-мотонейронов, f амма-мотонейроны активируют интрафузальные мышечные волокна, в результате чего активируются нервные окончания типа 1а и поток импульсов идет на альфа-мотонейроны или на вставочные мотонейроны, а от них к альфа-мотонейронам — это называется гамма-петля. Таким образом, гамма-мотонейроны выступают в роли вставочных нейронов, но с особым вариантом возбуждения: с участием периферического посредника в виде мышечных веретен. В спинном мозге имеются также нейроны, выполняющие роль пейсмекера. Они могут, автоматически возбуждаясь, активировать непосредственно альфа-мотонейроны без сигналов от супраслинальных двигательных систем. Однако у взрослого человека и даже у ребенка этот механизм активации альфа-мотонейронов, вероятно, полностью блокирован.
«Второй этаж» управления — это стволовые структуры: вестибулярные ядра, от которых идет вестибулоспинальный путь, красное ядро (руброспинальный путь), ретикулярная формация (ретикулоспинальный путь), покрышка четверохолмия (тектум, тектоспиналь- ный путь). Благодаря этим структурам регулируется мышечный тонус, поза как в условиях покоя, так и при выполнении целенаправленных движений. Этот «этаж» работает в тесном взаимодействии с мозжечком и корой мозга (экстрапирамидные пути, которые начинаются от клеток Беца двигательной коры мозга, обязательно связаны с соответствующими структурами ствола мозга).
«Третий этаж» — это кора. Зарождающийся в ассоциативных зонах коры замысел «поступает» в двигательную кору, откуда он направляется по пирамидному пути к альфа-мотонейронам спинного мозга (часть волокон через вставочные нейроны спинного мозга). Одновременно для коррекции движения и для того, чтобы данная фазная активность проходи-
ла в удобном положении, сигнал идет от клеток Беца к структурам ствола мозга и тем самым регулируется поза (экстрапирамидная система). Для того, чтобы движения были организованы правильно, выходящий из ассоциативной зоны коры «замысел» предварительно попадает к базальным ганглиям, где происходит коррекция и выбор программы действия» и возвращается к двигательной коре, откуда он идет по пирамидному пути. Параллельно, из ассоциативной коры сигнал попадает в мозжечок, а из него через таламус возвращается в двигательную кору (мозжечок также вносит свой вклад в составление программы, в коррекцию движения). Кольца (ассоциативная кора — базальные ганглии — таламус — двигательная кора, ассоциативная кора — мозжечок — таламус — двигательная кора) — тоже являются компонентом экстрапирамидной системы.
Все двигательные системы работают за счет обязательного использования сенсорной информации. Особая роль принадлежит здесь информации, идущей от рецепторов мышц (мышечные веретена, сухожильные рецепторы Гольджи, рецепторы суставов), от кожи (тактильные и болевые рецепторы), а также от вестибулярного анализатора, благодаря которому любое целенаправленное движение выполняется в удобный для организма позе вопреки воздействию сил земного притяжения.
РОЛЬ СПИННОГО МОЗГА В РЕГУЛЯЦИИ ДВИГАТЕЛЬНОЙ АКТИВНОСТИ
С участием спинного мозга осуществляются примитивные процессы регуляции деятель ности скелетных мышц, позволяющие выполнить фазные движения типа сгибания или раз гибания в соответствующих суставах, а также регулирующие тонус мышц. Регуляция тону са мышц осуществляется с участием двух видов рефлексов спинного мозга: миотатическил и позно-тонических. Фазная активность представлена сгибательными рефлексами и механизмами, инициирущими локомоторные движения (шаговые движения).
Миотатические рефлексы —это рефлексы, которые часто называют сухожильными, потому что в клинике для их выявления обычно производится удар неврологическим молоточком по сухожилию соответствующей мышцы. Эти рефлексы играют важную роль в поддержании тонуса мышцы, равновесия, они направлены против гравитационных сил. Например, когда человек занимает вертикальное положение, то за счет гравитации он может упасть (сгибание в коленном, голеностопном, тазобедренном суставах), но с участием миотатических рефлексов это не происходит, т. к. при растяжении мышцы активируются мышечные веретена, которые расположены параллельно экстрафузальным волокнам скелетной мышцы. Импульсация от этих рецепторов идет через афферентный нейрон и попадает на альфа- мотонейроны данной мышцы. В результате происходит укорочение экстрафузальных волокон. Тем самым длина мышцы возвращается к исходной. С позиций кибернетики этот рефлекс представляет пример регулирования по принципу рассогласования: если длина мышцы превышает заданную длину, то активируется управляющее устройство, которое вызывает изменение объекта управления — укорочение мышцы, в результате чего длина мышцы становится такой, как должна быть. Отметим, что импульсация от рецепторов одновременно через тормозные вставочные нейроны попадает на альфа-мотонейроны антагониста этой мышцы, поэтому при укорочении агониста мышца-антагонист не препятствует этому процессу.
Сигнал от супраспинальных механизмов, приходящий к альфа-мотонейронам, одновременно идет и на гамма-мотонейроны и тем самым возбуждает интрафузальные мышечные волокна: в этом случае даже в укороченной мышце веретена будут способны следить за длиной мышцы. Такое одновременное возбуждение альфа- и гамма-мотонейронов получило название альфа-гамма-коактивации.
О существовании механизма гамма-петли уже указывалось выше: ряд воздействий со стороны супраспинальных механизмов может идти не на вставочные мотонейроны и не на альфа-мотонейроны, а только на гамма-мотонейроны: их возбуждение приводит к натяжению ядерной сумки мышечного веретена и в результате этого даже в покоящейся нерастя -
нутой мышце возникает поток импульсов, идущий в спинной мозг, и, как и при миотатическом рефлексе, — возникает активация альфа-мотонейронов и сокращение данной мышцы.
Когда неврологическим молоточком наносится быстрый удар по сухожилию, то создаются условия для быстрого растяжения мышцы — возникает залп импульсов, который приводит к активации многих альфа-мотонейронов данной мышцы. Она сокращается, что ведет к разгибанию или сгибанию в соответствующем суставе. В неврологической практике сухожильные рефлексы используются для оценки состояния соответствующих сегментов спинного мозга. Например, коленный рефлекс отражает состояние 2—4 поясничных сегментов.
Сгибательныйрефлекс возникает под влиянием потока импульсов, идущих от кожных рецепторов — тактильных, температурных, болевых. Эти потоки называются афферентами сгибательного рефлекса. Все импульсы возбуждают альфа-мотонейроны сгибателя ипсилатеральной конечности и одновременно тормозят альфа-мотонейроны разгибателей данной конечности — происходит сгибание в соответствующем суставе, а в итоге — «уход» от повреждающего фактора. Этот рефлекс намного сложнее мяотатического, так как одновременно в силу дивергенции импульсация от афферентных нейронов распространяется на альфа-мотонейроны контрлатеральной конечности: активируются альфа-мотонейроны разгибателя и тормозятся альфа-мотонейроны сгибателей. В результате, контрлатеральная конечность выпрямляется, и тяжесть тела может быть перенесена на эту конечность (если она используется для реализации позы «стоя»). Таким образом, этот рефлекс демонстрирует координацию фазного движения с процессом сохранения позы (тонические рефлексы).
Если новорожденного поставить на твердую поверхность стола, поддерживая его, то можно отметить появление у него серии шагательных движений. Этот рефлекс называется рефлексом шагательных движений. Он сохраняется примерно до двух месяцев жизни. В этом рефлексе, однако, нет классической рефлекторной дуги и классического пути ее активации (рецептор — афферентный нейрон — эфферентный нейрон). Считается, что это один из вариантов функционирования нейронных образований, в которых заранее заложена «программа» действий. Эта программа весьма примитивна, но она существует и у ряда животных, зероятно, играет определенную роль (вспомним: петух, бегущий с отрезанной головой, а точнее — без головы). Поэтому этот «рефлекс» рассматривают как проявление комплекса фиксированных действий. У человека он уже к двум месяцам постнатального периода жизни подавляется супраспинальными механизмами и лишь при глубокой патологии ЦНС может проявиться вновь. Вероятно, для человека — это рудимент, поэтому он «уходит» из арсенала средств, используемых в ЦНС. Но другие комплексы фиксированных действий, например, лежащие в основе функций дыхательного центра, существуют.
Позно-тонические рефлексы спинного мозганаправлены на поддержание позы, с их помощью регулируется тонус мышц. Эти рефлексы возникают с проприорецепторов мышц шеи, рецепторов фасций шеи животного. Переключение с этих рецепторов осуществляется на уровне шейного отдела спинного мозга и приводит к изменению тонуса мышц при изменении положения головы и шеи. Позно-тонические рефлексы спинного мозга, вероятно, отчетливо выражены далеко не у всех млекопитающих, так как по мере усложнения организации мозга этот вид деятельности переходит к структурам ствола мозга. У лягушки при сохранении нейронов шейного отдела мозга возможно проявление позно-тонических рефлексов.
СПИНАЛЬНЫЙ ШОК. ПАТОЛОГИЧЕСКИЕ РЕФЛЕКСЫ
Спинальный шок развивается в результате прерывания супраспинальных влияний на мотонейроны спинного мозга. Происходит это при травме мозга. По неизвестным до сих пор причинам на определенное время деятельность нейронов спинного мозга приостанавливается. Чем выше развитие организма, тем длительнее период, в течение которого происходит восстановление функций спинного мозга. У лягушки этот процесс занимает минуты, у человека — недели и месяцы.
При нарушении супраспинальных влияний у человека может появиться группа спинальных рефлексов, которые имеются в норме лишь в первые дни и месяцы постнатального развития. Растормаживание этих примитивных рефлексов является клиническим признаком нарушений работъ1 мозга. Поэтому существуют определенные приемы, позволяющие выявить эти так называемые патологические рефлексы. Различают разгибательные и сгибательные патологические рефлексы: речь идет о тыльном разгибании большого пальца стопы (разгибательные рефлексы) и о подошвенном сгибании большого пальца стопы (сгибательные рефлексы). Например, разгибательный рефлекс Бабинского: при раздражении подошвы стопы штриховым движением от пятки до носка происходит разгибание большого пальца в сторону тыла. В норме этот рефлекс имеет место у детей (примерно до 2 лет), потом он тормозится и выявляется лишь при нарушении супраспинальных влияний. Рефлекс Оппенгейма — при надавливании на поверхность голени в области большеберцовой кости — от колена до голеностопного сустава — происходит тыльное разгибание большого пальца. Рефлекс Гордона — тыльное разгибание большого пальца происходит при сдавливании икроножной мышцы, рефлекс Шефера — то же самое происходит при сдавливании ахиллового сухожилия. Сгибательные рефлексы: рефлекс Россолимо — сгибание пальцев стопы и в том числе большого при быстром касательном ударе по подушечкам пальцев стопы, аналогично: подошвенное сгибание большого пальца при ударе молоточком по тылу стопы (рефлекс Бехтерева-Менделя), при ударе по подошвенной поверхности под пальцами стопы (рефлекс Жуковского), при ударе по пятке (рефлекс Бехтерева).
Имеются и патологические рефлексы, вызываемые при нанесении раздражителя на верхнюю конечность, например, сгибательный рефлекс Россолимо — сгибание пальцев кисти в ответ на быстрое касание пальцев, или рефлекс Жуковского — сгибание пальцев кисти при ударе молоточком по ладонной поверхности кисти, рефлекс Бехтерева — то же сгибание при ударе по тылу кисти.
К патологическим рефлексам относится хватательный рефлекс (Янушкевича) — он возникает при касании пальцев руки или стопы каким-либо предметом и проявляется в захватывании этого предмета, которое происходит непроизвольно: У детей этот рефлекс проявляется в возрасте до 2—4 месяцев. Иногда при этом удается приподнять ребенка над опорой: в этом случае хватательный рефлекс называют рефлексом Робинсона.
Итак, патологические рефлексы — это по сути истинные рефлексы, которые в процессе онтогенеза как рудиментарные рефлексы заторможены с помощью специальных механизмов, нарушаемых при заболевании мозга.
СТВОЛ МОЗГА И ЕГО РОЛЬ В РЕГУЛЯЦИИ ДВИГАТЕЛЬНЫХ ФУНКЦИЙ
Ствол мозга — это продолговатый мозг, мост (Варолиев мост) и средний мозг. Продолговатый мозг и мост вкупе называют задним мозгом.
Ствол мозга содержит важные структуры, принимающие участие в регуляции мышечной активности: двигательные ядра черепно-мозговых нервов, вестибулярные ядра, красное ядро, ретикулярную формацию, нейроны покрышки четверохолмия (тектум), а также черную субстанцию, о которой будет сказано в разделе «Базальные ганглии».
Как известно, часть черепно-мозговых нервов имеет двигательные ядра — скопление альфа-мотонейронов, которые принимают участие в регуляции сократительной активности поперечно-полосатых мышц. Так, Ш пара (глазодвигательный нерв) иннервирует 4 мышцы глазного яблока — внутреннюю, нижнюю и верхнюю прямые, нижнюю косую мышцы и мышцу, поднимающую верхнее веко. IV пара — блоковой нерв — иннервирует верхнюю косую мышцу глаза, VI пара — отводящий нерв — иннервирует наружную прямую мышцу глаза. V пара — тройничный нерв — иннервирует жевательную мускулатуру, VII пара — лицевой нерв — иннервирует всю мимическую мускулатуру, IX пара — языкоглоточный — иннервирует шилоглоточную мышцу, которая поднимает верхнюю часть глотки при глотании, X нерв — вагус — иннервирует мускулатуру глотки, мягкое небо, мышцы гортани,
надгортанник, верхнюю часть пищевода (здесь тоже имеется поперечно-полосатая мышца), XI нерв — добавочный — иннервирует голосовые мышцы, грудиноключичную мышцу, трапециевидную, а XII пара — подъязычный нерв — иннервирует мышцы языка и мышцы, двигающие язык вперед, вверх и вниз. Альфа-мотонейроны этих ядерных скоплений получают коллатерали от пирамидного пути (кортико-ядерный путь). При нарушении передачи возбуждения от коры до соответствующих мышц развивается паралич или парез. Знание анатомических и функциональных особенностей ядер черепно-мозговых нервов позволяет невропатологу поставить топический диагноз повреждений, возникающих в структурах мозга.
Ствол мозга принимает непосредственное участие в регуляции позы тела, используя для этих целей статические и стато-кинетические рефлексы. Эти рефлексы представляют собой механизмы перераспределения мышечного тонуса, в результате чего сохраняется удобная для животного (и человека) поза или происходит возвращение в эту позу из «неудобной» (соответственно — позно-тонические и выпрямительные рефлексы), а также сохраняется равновесие при ускорении (стато-кинетические рефлексы). В их реализации участвуют нейроны вестибулярных ядер, красного ядра и ретикулярной формации.
Вестибулярные ядра возбуждаются под влиянием адекватных раздражителей, действующих на вестибулярный аппарат. Одно из главных ядер — это ядро Дейтерса. От него начинается вестибулоспинальный путь, который осуществляет воздействие на альфа-мотонейроны спинного мозга. Нейроны вестибулярных ядер возбуждают альфа-мотонейроны разгибателей и одновременно по механизму реципрокной иннервации тормозят альфа-мотонейроны сгибателей. Благодаря этому при раздражении вестибулярного аппарата так меняется тонус верхних и нижних конечностей, что равновесие сохраняется. Вестибулярное ядро Дейтерса находится под контролем мозжечка. От вестибулярного аппарата к мозжечку идет прямой вестибулоцеребеллярный путь, т. е. мозжечок получает всю информацию от вестибулярного аппарата, Вместе с информацией, идущей от проприорецепторов и от рецепторов кожи, она перерабатывается в коре мозжечка (в основном — в архицеребеллуме) и поступает на ядро шатра мозжечка, откуда вновь идет на вестибулярные ядра, в том числе Дейтерса. Таким образом контролируется деятельность вестибулярных ядер. Не случайно, что патология мозжечка проявляется примерно теми же симптомами, что и патология вестибулярного аппарата и вестибулярных ядер.
Одновременно от вестибулярных ядер продолговатого мозга идет путь к так называемому медиальному продольному пучку. Этот пучок начинается от ядра Даркшевича и промежуточного ядра, находящихся в среднем мозге; пучок направлен в сторону спинного мозга и выполняет важную функцию — соединяет между собой в единый функциональный ансамбль все ядра нервов, участвующих в регуляции активности мышц глазного яблока (III, IV, VI пары). Благодаря этому движение обоих глазных яблок происходит синхронно и в норме у большинства людей отсутствует явление косоглазия. Сигналы, идущие от вестибулярных ядер, попадают на продольный медиальный пучок, благодаря чему при активации вестибулярного аппарата возникает явление нистагма глазных яблок: такие движения способствуют удержанию взора на точке фиксации (на сетчатку изображение падает примерно на одно и то же место, несмотря на то, что человек совершает движение).
Таким образом, при раздражении вестибулярного аппарата происходит перераспределение мышечного тонуса и изменение активности мышц глазного яблока, в результате чего человек (животное) способен сохранять равновесие и взор, несмотря на совершение движения или вопреки отклонению головы от обычного положения.
При патологии вестибулярного аппарата или вестибулярных ядер и вестибулоспинального тракта возникают такие симптомы как головокружение (это субъективное ощущение вращения предметов окружающего мира, которое возникает вследствие «неверной» информации, поступающей в ассоциативные участки коры), нарушение равновесия (атаксия статическая, атаксия стато-кинетическая) и возникающий спонтанно нистагм глазных яблок: горизонтальный, вертикальный или ротационный.

|
|
Было уже давно замечено, что при отделении красного ядра от нижележащих структур повышается тонус экстензоров. У кошки, например, это проявляется в резком выпрямлении передних и задних конечностей, выгибании спины и хвоста, это явление получило название — децеребрационная ригидность. Необычайность позы, возникающей в этих условиях, давно привлекала внимание исследователей, и, может, поэтому теперь известно много интересного о работе мозга. Именно анализ этого явления позволил установить, что нейроны красного ядра вызывают, в основном, возбуждение мышц- сгибателей. Поэтому при его нарушении преобладает влияние вестибулярного ядра (экстензорное) и возникает децеребрационная ригидность. Если вместе с перерезкой путей, идущих от красного ядра, нарушить вестибулярные ядра или вестибулоспинальный тракт, то ригидность исчезает. Итак, децеребрационная ригид-
ность — это феномен, позволивший в деталях разобраться в сложном устройстве двигательных механизмов ствола мозга.
Ретикулярная формация ствола мозга <— это структура, идущая в ростральном (к коре) направлении от спинного мозга к таламусу. Помимо участия в обработке сенсорной информации (неспецифический канал) ретикулярная формация выполняет и функции двигательной системы. Обнаружено, что имеется два скопления нейронов ретикулярной формации, причастных к этому: это нейроны продолговатого мозга и нейроны моста. Нейроны ретикулярной формации продолговатого мозга ведут себя точно так же, как нейроны красного ядра: они активируют альфа-мотонейроны сгибателей и тормозят альфа-мотонейроны разгибателей. Нейроны ретикулярной формации моста, наоборот, действуют, как нейроны вестибулярных ядер. Таким образом, можно условно все двигательные системы ствола мозга разделить на 2 класса (по их влиянию на мышцы): флексорные системы, повышающие активность сгибателей — это нейроны красного ядра и ретикулярной формации продолговатого мозга и 2) экстензорные системы — нейроны вестибулярных ядер и нейроны ретикулярной формации моста.
Ретикулярная формация, подобно вестибулярным ядрам и нейронам красного ядра, получает информацию от коры мозга (экстрапирамидный путь) и тесно связана с мозжечком: часть информации от мозжечка идет к нейронам продолговатого мозга (от ядер пробковидного и шаровидного мозжечка), а от ядра шатра — к нейронам, локализованным в мосту. Поэтому ретикулярная формация также принимает участие в регуляции позы. Вероятно, вследствие того, что ретикулярная формация — это коллектор неспецифического сенсорного потока, она, возможно, на основе этой информации участвует в регуляции мышечной активности. Пока, однако, непонятна необходимость в нейронах ретикулярной формации, необходимость дублирования нейронов вестибулярных ядер и красного ядра.
Итак, ствол мозга обеспечивает поддержание равновесия в условиях покоя и ускорения. Это проявляется в наличии статических и стато-кинетических рефлексов. Впервые их подробно исследовал голландский физиолог Р. Магнус, поэтому статические рефлексы называют «рефлексы Магнуса».
Статические рефлексы условно делят на позно-тонические и выпрямительные. Оба вида рефлексов возникают в результате раздражения рецепторов вестибулярного аппарата, про- приорецепторов мышц и рецепторов фасции шеи, а также (выпрямительные) — при активации рецепторов кожи. Основная структура, участвующая в реализации этих рефлексов — вестибулярные ядра. Красное ядро и ретикулярная формация также причастны к этим рефлексам. Позно-тонические рефлексы, возникающие с шейных мышц, иногда называют шейно-тоническими, а возникающие с вестибулярного аппарата — вестибуло-тонически- ми. Но, учитывая, что в норме одновременно возбуждаются рецепторы мышц шеи и вестибулярный аппарат, целесообразно говорить о позно-тонических рефлексах. У животного и человека эти рефлексы хорошо выявляются в раннем возрасте. Например, у грудных детей наблюдается лабиринтный тонический рефлекс: у ребенка, лежащего на спине, повышен тонус разгибателей шеи, спины, ног Если же перевернуть его на живот, то увеличивается тонус сгибателей шеи, спины, конечностей. У них же имеет место симметричный шейный тонический рефлекс: при пассивном сгибании головы ребенка, лежащего на спине, происходит повышение тонуса сгибателей рук и повышение тонуса разгибателей ног. При разгибании головы наблюдаются противоположные процессы. Асимметричный шейный тонический рефлекс: голову ребенка, лежащего на спине, поворачивают в сторону так, чтобы подбородок касался плеча, при этом у ребенка снижается тонус сгибателей конечностей (к которым обращено лицо) и даже возрастает тонус разгибателей, а на противоположной стороне на руке и ноге возрастает тонус сгибателей. Все эти рефлексы у детей тормозятся примерно к 2—3 месяцам.
У животных позно-тонические рефлексы легко наблюдать при нарушении связей ствола мозга с передним мозгом. Например, кошка пытается поймать колбаску — голова ее приподнята. В этом случае тонус мышц-разгибателей передних конечностей увеличен, а на
задних конечностях повышен тонус мышц-сгибателей. Кошка «пьет» молоко — все наоборот. Поворот головы кошки вправо вызывает разгибание правых конечностей, а поворот головы влево — разгибание левых конечностей. Это совпадает с асимметричным шейным тоническим рефлексом новорожденного ребенка.
При наличии выраженных супраспинальных и супрастволовых влияний в «чистом» виде эти рефлексы у взрослого выявить трудно. Но при выполнении быстрых и резких движений можно выявить все эти рефлексы.
Итак, позно-топические рефлексы направлены на поддержание равновесия тела в условиях, когда смещается голова и шея.
Выпрямительные рефлексы проявляются в том, что животное из «непривычного», несвойственного для него положения переходит в естественное положение. Таким рефлексом является феномен приземления кошки на 4 лапы, независимо от того, как ее подбрасывают.
Стато-кннетические рефлексы направлены на сохранение позы (равновесия) и ориентации в пространстве при изменении скорости движения (наличии угловых и вертикальных ускорений). В частности, таким рефлексом является упомянутый выше нистагм глазных яблок и головы: это медленное вращение в сторону, противоположную вращению, и быстрый возврат в сторону вращения. Сигнал идет от вестибулярного аппарата через вестибулярные ядра на медиальный продольный пучок по направлению к ядрам III,IV и VI пары и приводит к изменению положения глазных яблок. Этим сохраняется ориентация в пространстве. Одновременно через вестибулоспинальный тракт меняется тонус мышц конечностей и туловища, в результате чего сохраняется равновесие.
К стато-кинетическим рефлексам относят лифтные рефлексы: при подъеме вверх повышается тонус сгибателей ног, а при опускании вниз возрастает тонус разгибателей.
Среди рефлексов ствола мозга необходимо отметить сторожевой и ориентировочный рефлексы. Они осуществляются с участием нейронов тектума: в ответ на зрительный сигнал (световой) или на звуковой происходит активация соответствующих рецепторов, сигнал достигает первичных центров (зрительные — передние или верхние бугры четверохолмия, слуховые — задние или нижние бугры), переключается на тектоспинальный тракт — возникают ориентировочные рефлексы: поворот головы в ту сторону, откуда поступает сигнал, и одновременно происходит перераспределение тонуса мышцы, в том числе за счет изменения положения головы, благодаря которому сохраняется равновесие и возникают благоприятные условия для совершения нападения на «врага» или убегания.
С участием ядер V, IX, X XI, XII пар нервов осуществляются произвольные движения типа жевания (альфа-мотонейроны V пары) и глотания (последовательное возбуждение альфа-мотонейронов языкоглоточного, блуждающего, добавочного и подъязычного нервов). При повреждении этих структур нарушаются акты жевания и глотания. С помощью нейронов IX и X пары идет управление голосовыми связками, а с помощью нейронов XII пары — языком. Поэтому при повреждении этих структур нарушается фонация и артикуляция (речь).
ФИЗИОЛОГИЯ МОЗЖЕЧКА, ЕГО ВЛИЯНИЕ НА МОТОРНЫЕ ФУНКЦИИ
У мозжечка имеется кора. Она содержит 3 слоя — молекулярный, ганглиозный (слой Пуркинье) и гранулярный (зернистый). Белое вещество мозжечка содержит ядро шатра, пробковидное и шаровидное ядра, зубчатое ядро. Принцип работы мозжечка заключается в следующем: поступает обширная информация, в том числе от вестибулярного аппарата, от мышечных рецепторов, от кожных рецепторов, а также от коры больших полушарий. Эта информация обрабатывается в коре мозжечка. Результаты обработки подаются на ядра мозжечка, которые управляют деятельностью красного ядра, вестибулярного ядра, ретикулярной формации. Кроме того, информация идет в кору больших полушарий, где используется для составления точных программ выполнения сложных движений.
Кора мозжечка представлена 6 типами клеток: 1) клетки-зерна (расположены в зернистом, самом нижнем слое коры мозжечка), они являются возбуждающими в отличие от ос-
 |
тальных 5 видов клеток, которые выполняют роль тормозных структур; 2) клетки Гольджи (тормозные, расположены в 3-м слое); 3) клетки Пуркинье или грушевидные клетки — самые большие клетки мозжечка, тормозные, их аксоны — это единственный выход из коры
В мозжечок поступает 2 типа волокон — лиановидные или лазящие и моховидные. Лазящие волокна непосредственно контактируют с телом или дендритами клеток Пуркинье, возбуждают эти клетки и тем самым усиливают тормозное влияние клеток Пуркинье на ядра мозжечка. По данным ряда авторов, лазящие волокна несут информацию от рецепторов мышц, суставов, от вестибулярных ядер (через нейроны нижней оливы). По моховидным волокнам информация поступает тоже от рецепторов мышц, сухожилий, вестибулярного аппарата, но идет при этом от соответствующих рецепторов, не поступая в нейроны оливы. Моховидные волокна передают свою информацию на клетки-зерна, которые в свою очередь передают ее на корзинчатые, звездчатые и другие клетки, возбуждая их, что приводит к торможению активности клеток Пуркинье (снятию тормозного влияния коры мозжечка на ядра мозжечка).
Таким образом, путь, идущий на клетки Пуркинье через лазящие волокна, способствует усилению тормозного влияния клеток Пуркинье на ядра мозжечка, а путь, идущий через моховидные волокна, наоборот, снимает это тормозное влияние. Очевидно, в этом и состоит смысл разделения информации от одних и тех же рецепторов на два потока.
С точки зрения функций, которые выполняет мозжечок, многие исследователи делят его на 3 части. Принцип деления у различных авторов свой. Наиболее распространенным является деление мозжечка на 3 части: архиоцеребеллум (древний мозжечок или вестибулоцеребел- лум), палеоцеребеллум (старый мозжечок) и неоцеребеллум (новый мозжечок). Р. Шмидт и Г. Тевс (1996), например, придерживаются иного деления: они предлагают разделить мозжечок продольными линиями на три части: внутреннюю (червь мозжечка), среднюю и латеральную; это примерно соответствует по функции делению на архио-, палео- и неоцеребеллум. Считается, что кора архиоцеребеллума (или внутренняя" часть, по Р. Шмидту и Г. Тевсу, 1996) связана с ядром шатра. Это ядро регулирует активность вестибулярных полагают, что флокулонодулярная часть мозжечка, вестибулярным регулятором: изменяя активность нейронов вестибулярных ядер, мозжечок тем самым влияет на процессы равновесия и все, что связано с деятельностью вестибулярных ядер. Ядро шатра влияет и на нейроны ретикулярной формации моста.
Функция палеоцеребеллума, или средней части коры (по Р. Шмидту и Г. Тевсу, 1996), — это взаимная координация позы и целенаправленного движения, а также коррекция выполнения сравнительно медленных движений на основе механизма обратной связи. Эта функция реализуется с участием двух ядер мозжечка — пробковидного и шаровидного (промежуточных ядер). Они влияют на деятельность красного ядра и Рис 22 Безмозжечковое животноеРетикУляРн°й формации, расположенной в Рас<:тройсгва;обнару«ивающиесяТпри' ”Р™о”говато“. Р™ 'исть "°зже™ ходьбе: атаксия, астения, астазия и т. д. работает на основании информации от мы-
шечных рецепторов и двигательной коры. Функция коррекции медленных движений имеет важное значение в процессе обучения, но она не может использоваться при выполнении быстрых и очень сложных движений (имеет место дефицит времени).
Неоцеребеллум (или по Р.. Шмидту и П Тевсу, 1996), латеральная часть мозжечка — играет важную роль в программировании сложных движений, выполнение которых идет без использования механизма обратных связей. Информация в неоцеребеллум поступает от ассоциативных зон коры («замысел»), которая вначале доставляется в нейроны моста (понтийный центр), откуда по понтоцеребеллярному пути она поступает в неоцеребеллум. От нейронов коры мозжечка информация идет на зубчатое ядро, которое у приматов достигает огромных размеров. От этого ядра информация идет через таламус к двигательной коре, откуда она поступает по пирамидному и экстрапирамидному пути (в том числе к красному ядру, вестибулярному ядру, чтобы сохранить равновесие при выполнении быстрого движения) к альфа-мотонейронам спинного мозга. В итоге возникает целенаправленное движение, выполняемое с большой скоростью, например, игра на фортепиано.
Ядра мозжечка находятся под влиянием нейронов Пуркинье. Когда активность этих тормозных нейронов возрастает, влияние ядер мозжечка на стволовые структуры (вестибулярное ядро, красное ядро) уменьшается. Когда активность нейронов Пуркинье снижается, снимается их тормозное действие на ядра мозжечка и тем самым мозжечок более активно влияет на функции стволовых структур.
РОЛЬ НАЗАЛЬНЫХ ЯДЕР В ФОРМИРОВАНИИ ТОНУСА И ЛОКОМОЦИЙ
Базальные ганглии (ядра) — это структуры переднего мозга: хвостатое ядро, скорлупа чечевицеобразного тела или просто скорлупа, бледный шар, субталамическое ядро; функционально к этим структурам относят черную субстанцию среднего мозга. Функция этих структур до настоящего времени изучена недостаточно полно. Считается, что базальные ганглии, как и мозжечок, используются в качестве системы, в которой уточняется программа выполнения сложных движений. Полагают, что информация от ассоциативных участков коры, т. е. мест, где зарождается замысел движения, поступает одновременно к мозжечку и параллельно к базальным ганглиям, а от них через таламус поступает в двигательную кору, где уточненная программа используется для управления: информация идет через пирамидный и экстрапирамидный путь к альфа-мотонейронам спинного мозга. При нарушении этой сложной связи пр
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|