- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Соматосенсорная система 1 страница
сенсорных систем, которые пока недоступны для экспериментальных методов.
Протезирование сенсорных функций практически проверяет истинность наших знаний о них. Примером могут быть электро-фосфеновые зрительные протезы, которые восстанавливают зрительное восприятие у слепых людей разными сочетаниями точечных электрических раздражений зрительной области коры большого мозга.
14.1.2. Общие принципы строения сенсорных систем
Основными общими принципами построения сенсорных систем высших позвоночных животных и человека являются следующие:
1) многослойность, т. е. наличие нескольких слоев нервных клеток, первый из которых связан с рецепторами, а последний — с нейронами моторных областей коры большого мозга. Это свойство дает возможность специализировать нейронные слои на переработке разных видов сенсорной информации, что позволяет организму быстро реагировать на простые сигналы, анализируемые уже на первых уровнях сенсорной системы. Создаются также условия для избирательного регулирования свойств нейронных слоев путем восходящих влияний из других отделов мозга;
2) многоканальность сенсорной системы, т. е. наличие в каждом слое множества (от десятков тысяч до миллионов) нервных клеток, связанных с множеством клеток следующего слоя. Наличие множества таких параллельных каналов обработки и передачи информации обеспечивает сенсорной системе точность и детальность анализа сигналов и большую надежность;
3) разное число элементов в соседних слоях, что формирует «сенсорные воронки». Так, в сетчатке глаза человека насчитывается 130 млн фоторецепторов, а в слое ганглиозных клеток сетчатки нейронов в 100 раз меньше («суживающаяся воронка»). На следующих уровнях зрительной системы формируется «расширяющаяся воронка»: число нейронов в первичной проекционной области зрительной области коры в тысячи раз больше, чем ганглиозных клеток сетчатки. В слуховой и в ряде других сенсорных систем от рецепторов к коре большого мозга идет «расширяющаяся воронка». Физиологический смысл «суживающейся воронки» заключается в уменьшении избыточности информации, а «расширяющейся» — в обеспечении дробного и сложного анализа разных признаков сигнала;
4) дифференциация сенсорной системы по вертикали и по горизонтали. Дифференциация по вертикали заключается в образовании отделов, каждый из которых состоит из нескольких нейронных слоев. Таким образом, отдел представляет собой более крупное морфофункциональное образование, чем слой нейронов. Каждый отдел (например, обонятельные луковицы, кохлеарные ядра слуховой системы или коленчатые тела) осуществляет определенную функцию.
Дифференциация по горизонтали заключается в различных свойствах рецепторов, нейронов и связей между ними в пределах каждого из слоев. Так, в зрении работают два параллельных нейронных канала, идущих от фоторецепторов к коре большого мозга и по-разному перерабатывающих информацию, поступающую от центра и от периферии сетчатки глаза.
14.1.3. Основные функции сенсорной системы
Сенсорная система выполняет следующие основные функции, или операции, с сигналами: 1) обнаружение; 2) различение; 3) передачу и преобразование; 4) кодирование; 5) детектирование признаков; 6) опознание образов. Обнаружение и первичное различение сигналов
обеспечивается рецепторами, а детектирование и опознание сигналов — нейронами коры больших полушарий. Передачу, преобразование и кодирование сигналов осуществляют нейроны всех слоев сенсорных систем.
Обнаружение сигналов. Оно начинается в рецепторе — специализированной клетке, эволюционно приспособленной к восприятию раздражителя определенной модальности из внешней или внутренней среды и преобразованию его из физической или химической формы в форму нервного возбуждения.
Классификация рецепторов. В практическом отношении наиболее важное значение имеет психофизиологическая классификация рецепторов по характеру ощущений, возникающих при их раздражении. Согласно этой классификации, у человека различают зрительные, слуховые, обонятельные, вкусовые, осязательные рецепторы, термо-, проприо- и вестибулорецепторы (рецепторы положения тела и его частей в пространстве) и рецепторы боли.
Существуют рецепторы внешние (экстерорецепторы) и внутренние (интерорецепторы). К экстерорецепторам относятся слуховые, зрительные, обонятельные, вкусовые, осязательные. К интерорецепторам относятся вестибуло- и проприорецепторы (рецепторы опорно-двигательного аппарата), а также висцерорецепторы (сигнализирующие о состоянии внутренних органов).
По характеру контакта со средой рецепторы делятся на дистантные, получающие информацию на расстоянии от источника раздражения (зрительные, слуховые и обонятельные), и контактные — возбуждающиеся при непосредственном соприкосновении с раздражителем (вкусовые, тактильные).
В зависимости от природы раздражителя, на который они оптимально настроены, рецепторы могут быть разделены на фоторецепторы, механорецепторы, к которым относятся слуховые, вестибулярные рецепторы, и тактильные рецепторы кожи, рецепторы опорно-двигательного аппарата, барорецепторы сердечно-сосудистой системы; хеморецепторы, включающие рецепторы вкуса и обоняния, сосудистые и тканевые рецепторы; терморецепторы (кожи и внутренних органов, а также центральные термочувствительные нейроны); болевые (ноцицептивные) рецепторы.
Все рецепторы делятся на первично-чувствующие и вторично-чувствующие. К первым относятся рецепторы обоняния, тактильные и проприорецепторы. Они различаются тем, что преобразование энергии раздражения в энергию нервного импульса происходит у них в первом нейроне сенсорной системы. К вторично-чувствующим относятся рецепторы вкуса, зрения, слуха, вестибулярного аппарата. У них между раздражителем и первым нейроном находится специализированная рецепторная клетка, не генерирующая импульсы. Таким образом, первый нейрон возбуждается не непосредственно, а через рецепторную (не нервную) клетку.
Общие механизмы возбуждения рецепторов. При действии стимула на рецепторную клетку происходит преобразование энергии внешнего раздражения в рецепторный сигнал, или трансдукция сенсорного сигнала. Этот процесс включает в себя три основных этапа: 1) взаимодействие стимула, т. е. молекулы пахучего или вкусового вещества (обоняние, вкус), кванта света (зрение) или механической силы (слух, осязание) с рецепторной белковой молекулой, которая находится в составе клеточной мембраны рецепторной клетки; 2) внутриклеточные процессы усиления и передачи сенсорного стимула в пределах рецепторной клетки; и 3) открывание находящихся в мембране рецептора ионных каналов, через которые начинает течь ионный ток, что, как правило, приводит к деполяризации клеточной мембраны рецепторной клетки (возникновению так называемого рецепторного потенциала). В первично-чувствующих рецепторах этот потенциал действует на наиболее чувствительные участки мембраны, способные генерировать потенциалы действия — электрические нервные импульсы. Во вторично-чувствующих рецепторах рецепторный потенциал вызывает выделение квантов медиатора из пресинаптического окончания рецепторной клетки. Медиатор (например, ацетилхолин), воздействуя на постсинаптическую мембрану первого нейрона, изменяет ее поляризацию (генерируется постсинаптический потенцил). Постсинаптический потенциал первого нейрона сенсорной системы называют генераторным потенциалом, так как он вызывает генерацию импульсного ответа. В первично-чувствующих рецепторах рецепторный и генераторный потенциалы — одно и то же.
Абсолютную чувствительность сенсорной системы измеряют порогом реакции. Чувствительность и порог — обратные понятия: чем выше порог, тем ниже чувствительность, и наоборот. Обычно принимают за пороговую такую силу стимула, вероятность восприятия которого равна 0,5 или 0,75 (правильный ответ о наличии стимула в половине или в 3/4 случаев его действия). Более низкие значения интенсивности считаются подпороговыми, а более высокие — надпороговыми. Оказалось, что и в подпороговом диапазоне реакция на сверхслабые раздражители возможна, но она неосознаваема (не доходит до порога ощущения). Так, если снизить интенсивность вспышки света настолько, что человек уже не может сказать, видел он ее или нет, от его руки можно зарегистрировать неощущаемую кожно-гальваническую реакцию на данный сигнал.
Чувствительность рецепторных элементов к адекватным раздражителям, к восприятию которых они эволюционно приспособлены, предельно высока. Так, обонятельный рецептор может возбудиться при действии одиночной молекулы пахучего вещества, фоторецептор — одиночным квантом света. Чувствительность слуховых рецепторов также предельна: если бы она была выше, мы слышали бы постоянный шум из-за теплового движения молекул.
Различение сигналов. Важная характеристика сенсорной системы — способность замечать различия в свойствах одновременно или последовательно действующих раздражителей. Различение начинается в рецепторах, но в этом процессе участвуют нейроны всей сенсорной системы. Оно характеризует то минимальное различие между стимулами, которое сенсорная система может заметить (дифференциальный, или разностный, порог).
Порог различения интенсивности раздражителя практически всегда выше ранее действовавшего раздражения на определенную долю (закон Вебера). Так, усиление давления на кожу руки ощущается, если увеличить груз на 3% (к 100-граммовой гирьке надо добавить 3 г, а к 200-граммовой — 6 г). Эта зависимость выражается формулой: dI/I= const, где I — сила раздражения, dI — ее едва ощущаемый прирост (порог различения), const — постоянная величина (константа). Аналогичные соотношения получены для зрения, слуха и других органов чувств человека.
Зависимость силы ощущения от силы раздражения (закон Вебера—Фехнера) выражается формулой: E=a-logI+b, где Е — величина ощущения, I — сила раздражения, а и b — константы, различные для разных модальностей стимулов. Согласно этой формуле, ощущение увеличивается пропорционально логарифму интенсивности раздражения.
Выше упоминалось о различении силы раздражителей. Пространственное различение основано на распределении возбуждения в слое рецепторов и в нейронных слоях. Так, если два раздражителя возбудили два соседних рецептора, то различение этих раздражителей невозможно и они будут восприняты как единое целое. Необходимо, чтобы между двумя возбужденными рецепторами находился хотя бы один невозбужденный. Для временного различения двух раздражений необходимо, чтобы вызванные ими нервные процессы не сливались во времени и чтобы сигнал, вызванный вторым стимулом, не попадал в рефрактерный период от предыдущего раздражения.
Передача и преобразование сигналов. Процессы преобразования и передачи сигналов в сенсорной системе доносят до высших центров мозга наиболее важную (существенную) информацию о раздражителе в форме, удобной для его надежного и быстрого анализа.
Преобразования сигналов могут быть условно разделены на пространственные и временные. Среди пространственных преобразований выделяют изменения соотношения разных частей сигнала. Так, в зрительной и соматосенсорной системах на корковом уровне значительно искажаются геометрические пропорции представительства отдельных частей тела или частей поля зрения. В зрительной области коры резко расширено представительство информационно наиболее важной центральной ямки сетчатки при относительном сжатии проекции периферии поля зрения («циклопический глаз»). В соматосенсорной области коры также преимущественно представлены наиболее важные для тонкого различения и организации поведения зоны — кожа пальцев рук и лица («сенсорный гомункулюс»).
Для временных преобразований информации во всех сенсорных системах типично сжатие, временная компрессия сигналов: переход от длительной (тонической) импульсации нейронов на нижних уровнях к коротким (фазическим) разрядам нейронов высоких уровней.
Ограничение избыточности информации и выделение существенных признаков сигналов. Зрительная информация, идущая от фоторецепторов, могла бы очень быстро насытить все информационные резервы мозга. Избыточность сенсорных сообщений ограничивается путем подавления информации о менее существенных сигналах. Менее важно во внешней среде то, что неизменно либо изменяется медленно во времени и в пространстве. Например, на сетчатку глаза длительно действует большое световое пятно. Чтобы не передавать все время в мозг информацию от всех возбужденных рецепторов, сенсорная система пропускает в мозг сигналы только о начале, а затем о конце раздражения, причем до коры доходят сообщения только от рецепторов, которые лежат по контуру возбужденной области.
Кодирование информации. Кодированием называют совершаемое по определенным правилам преобразование информации в условную форму — код. В сенсорной системе сигналы кодируются двоичным кодом, т. е. наличием или отсутствием электрического импульса в тот или иной момент времени. Такой способ кодирования крайне прост и устойчив к помехам. Информация о раздражении и его параметрах передается в виде отдельных импульсов, а также групп или «пачек» импульсов («залпов» импульсов). Амплитуда, длительность и форма каждого импульса одинаковы, но число импульсов в пачке, частота их следования, длительность пачек и интервалов между ними, а также временной «рисунок» пачки различны и зависят от характеристик стимула. Сенсорная информация кодируется также числом одновременно возбужденных нейронов, а также местом возбуждения в нейронном слое.
Особенности кодирования в сенсорных системах. В отличие от телефонных или телевизионных кодов, которые декодируются восстановлением первоначального сообщения в исходном виде, в сенсорной системе такого декодирования не происходит. Еще одна важная особенность нервного кодирования — множественность и перекрытие кодов. Так, для одного и того же свойства сигнала (например, его интенсивности) сенсорная система использует несколько кодов: частотой и числом импульсов в пачке, числом возбужденных нейронов и их локализацией в слое. В коре большого мозга сигналы кодируются последовательностью включения параллельно работающих нейронных каналов, синхронностью ритмических импульсных разрядов, изменением их числа. В коре используется также позиционное кодирование. Оно заключается в том, что какой-то признак раздражителя вызывает возбуждение определенного нейрона или небольшой группы нейронов, расположенных в определенном месте нейронного слоя. Например, возбуждение небольшой локальной группы нейронов зрительной области коры означает, что в определенной части поля зрения появилась световая полоска определенного размера и ориентации.
Для периферических отделов сенсорной системы типично временное кодирование признаков раздражителя, а на высших уровнях происходит переход к преимущественно пространственному (в основном позиционному) коду.
Детектирование сигналов. Это избирательное выделение сенсорным нейроном того или иного признака раздражителя, имеющего поведенческое значение. Такой анализ осуществляют нейроны-детекторы, избирательно реагирующие лишь на определенные параметры стимула. Так, типичный нейрон зрительной области коры отвечает разрядом лишь на одну определенную ориентацию темной или светлой полоски, расположенной в определенной части поля зрения. При других наклонах той же полоски ответят другие нейроны. В высших отделах сенсорной системы сконцентрированы детекторы сложных признаков и целых образов. Примером могут служить детекторы лица, найденные недавно в нижневисочной области коры обезьян (предсказанные много лет назад, они были названы «детекторы моей бабушки»). Многие детекторы формируются в онтогенезе под влиянием окружающей среды, а у части из них детекторные свойства заданы генетически.
Опознание образов. Это конечная и наиболее сложная опера-ля сенсорной системы. Она заключается в отнесении образа к эму или иному классу объектов, с которыми ранее встречался организм, т. е. в классификации образов. Синтезируя сигналы от нейронов-детекторов, высший отдел сенсорной системы формирует «образ» раздражителя и сравнивает его с множеством образов, хранящихся в памяти. Опознание завершается принятием решения о том, с каким объектом или ситуацией встретился организм. В результате этого происходит восприятие, т. е. мы осознаем, чье лицо видим перед собой, кого слышим, какой запах чувствуем.
Опознание часто происходит независимо от изменчивости сигнала. Мы надежно опознаем, например, предметы при различной их освещенности, окраске, размере, ракурсе, ориентации и положении в поле зрения. Это означает, что сенсорная система формирует независимый от изменений ряда признаков сигнала (инвариантный) сенсорный образ.
Переработку информации в сенсорной системе осуществляют процессы возбудительного и тормозного межнейронного взаимодействия. Возбудительное взаимодействие заключается в том, что аксон каждого нейрона, приходя в вышележащий слой сенсорной системы, контактирует с несколькими нейронами, каждый из которых получает сигналы от нескольких клеток предыдущего слоя.
Совокупность рецепторов, сигналы которых поступают на данный нейрон, называют его рецептивным полем. Рецептивные поля соседних нейронов частично перекрываются (рис. 14.1). В результате такой организации связей в сенсорной системе образуется так называемая нервная сеть. Благодаря ей повышается чувствительность системы к слабым сигналам, а также обеспечивается высокая приспособляемость к меняющимся условиям среды.
Тормозная переработка сенсорной информации основана на том, что обычно каждый возбужденный сенсорный нейрон активирует тормозный интернейрон. Интернейрон в свою очередь подавляет импульсацию как самого возбудившего его элемента (последовательное, или возвратное, торможение), так и его соседей по слою (боковое, или латеральное, торможение). Сила этого торможения тем больше, чем сильнее возбужден первый элемент и чем ближе к нему соседняя клетка. Значительная часть операций по снижению избыточности и выделению наиболее существенных сведений о раздражителе производится латеральным торможением.
14.1.5. Адаптация сенсорной системы
Сенсорная система обладает способностью приспосабливать свои свойства к условиям среды и потребностям организма. Сенсорная адаптация — общее свойство сенсорных систем, заключающееся в приспособлении к длительно действующему (фоновому) раздражителю. Адаптация проявляется в снижении абсолютной и повышении дифференциальной чувствительности сенсорной системы. Субъективно адаптация проявляется в привыкании к действию постоянного раздражителя (например, мы не замечаем непрерывного давления на кожу привычной одежды).
Адаптационные процессы начинаются на уровне рецепторов, охватывая и все нейронные уровни сенсорной системы. Адаптация слаба только в вестибуло- и проприорецепторах. По скорости данного процесса все рецепторы делятся на быстро- и медленно адаптирующиеся. Первые после развития адаптации практически не посылают в мозг информации о длящемся раздражении. Вторые эту информацию передают в значительно ослабленном виде. Когда действие постоянного раздражителя прекращается, абсолютная чувствительность сенсорной системы восстанавливается. Так, в темноте абсолютная чувствительность зрения резко повышается.
В сенсорной адаптации важную роль играет эфферентная регуляция свойств сенсорной системы. Она осуществляется за счет нисходящих влияний более высоких на более низкие ее отделы. Происходит как бы перенастройка свойств нейронов на оптимальное восприятие внешних сигналов в изменившихся условиях. Состояние разных уровней сенсорной системы контролируется также ретикулярной формацией, включающей их в единую систему, интегрированную с другими отделами мозга и организма в целом. Эфферентные влияния в сенсорных системах чаще всего имеют тормозной характер, т. е. приводят к уменьшению их чувствительности и ограничивают поток афферентных сигналов.
Общее число эфферентных нервных волокон, приходящих к рецепторам или элементам какого-либо нейронного слоя сенсорной системы, как правило, во много раз меньше числа афферентных нейронов, приходящих к тому же слою. Это определяет важную особенность эфферентного контроля в сенсорных системах: его широкий и диффузный характер. Речь идет об общем снижении чувствительности значительной части нижележащего нейронного слоя.
14.1.6. Взаимодействие сенсорных систем
Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и корковом уровнях. Особенно широка интеграция сигналов в ретикулярной формации. В коре большого мозга происходит интеграция сигналов высшего порядка. В результате образования множественных связей с другими сенсорными и неспецифическими системами многие корковые нейроны приобретают способность отвечать на сложные комбинации сигналов разной модальности. Это особенно свойственно нервным клеткам ассоциативных областей коры больших полушарий, которые обладают высокой пластичностью, что обеспечивает перестройку их свойств в процессе непрерывного обучения опознанию новых раздражителей. Межсенсорное (кросс-модальное) взаимодействие на корковом уровне создает условия для формирования «схемы (или карты) мира» и непрерывной увязки, координации с ней собственной «схемы тела» организма.
14.2. ЧАСТНАЯ ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ 14.2.1. Зрительная система
Зрение эволюционно приспособлено к восприятию электромагнитных излучений в определенной, весьма узкой части их диапазона (видимый свет). Зрительная система дает мозгу более 90% сенсорной информации. Зрение — многозвеньевой процесс, начинающийся с проекции изображения на сетчатку уникального периферического оптического прибора — глаза. Затем происходят возбуждение фоторецепторов, передача и преобразование зрительной информации в нейронных слоях зрительной системы, а заканчивается зрительное восприятие принятием высшими корковыми отделами этой системы решения о зрительном образе.
Строение и функции оптического аппарата глаза. Глазное яблоко имеет шарообразную форму, что облегчает его повороты для наведения на рассматриваемый объект. На пути к светочувствительной оболочке глаза (сетчатке) лучи света проходят через несколько прозрачных сред — роговицу, хрусталик и стекловидное тело. Определенная кривизна и показатель преломления роговицы и в меньшей мере хрусталика определяют преломление световых лучей внутри глаза (рис. 14.2).
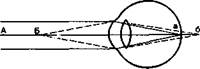
Преломляющую силу любой оптической системы выражают в диоптриях (D). Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила здорового глаза составляет 59D при рассматривании далеких и 70,5D — при рассматривании близких предметов. Чтобы схематически представить проекцию изображения предмета на сетчатку, нужно провести линии от его концов через узловую точку (в 7 мм сзади от роговой оболочки). На сетчатке получается изображение, резко уменьшенное и перевернутое вверх ногами и справа налево (рис. 14.3).
Аккомодация. Аккомодацией называют приспособление глаза к ясному видению объектов, удаленных на разное расстояние. Для ясного видения объекта необходимо, чтобы он был сфокусирован на сетчатке, т. е. чтобы лучи от всех точек его поверхности проецировались на поверхность сетчатки (рис. 14.4). Когда мы смотрим на далекие предметы (А), их изображение (а) сфокусировано на сетчатке и они видны ясно. Зато изображение (б) близких предметов (Б) при этом расплывчато, так как лучи от них собираются за сетчаткой. Главную роль в аккомодации играет хрусталик, изменяющий свою кривизну и, следовательно, преломляющую способность. При рассматривании близких предметов хрусталик делается более выпуклым (см. рис. 14.2), благодаря чему лучи, расходящиеся от какой-либо точки объекта, сходятся на сетчатке. Механизмом аккомодации является сокращение ресничных мышц, которые изменяют выпуклость хрусталика. Хрусталик заключен в тонкую прозрачную капсулу, которую всегда растягивают, т. е. уплощают, волокна ресничного пояска (циннова связка). Сокращение гладких мышечных клеток ресничного тела уменьшает тягу цинновых связок, что увеличивает выпуклость хрусталика в силу его эластичности. Ресничные мышцы иннервируются парасимпатическими волокнами глазодвигательного нерва. Введение в глаз атропина вызывает нарушение передачи возбуждения к этой мышце, ограничивает аккомодацию глаза при рассматривании близких предметов. Наоборот, парасимпатомиметические вещества — пилокарпин и эзерин — вызывают сокращение этой мышцы.
Для нормального глаза молодого человека дальняя точка ясного видения лежит в бесконечности. Далекие предметы он рассматривает без всякого напряжения аккомодации, т. е. без сокращения
Рис. 14.4. Ход лучей от близкой и далекой точек. Объяснение в тексте.
| 211 |
|
|
| Рис. 14.5. Рефракция в нормальном (А), близоруком (Б) и дальнозорком (Г) глазу и оптическая коррекция близорукости (В) и дальнозоркости (Д) (схема). |
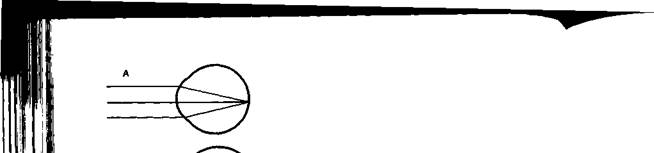 ресничной мышцы. Ближайшая точка ясного видения находится на расстоянии 10 см от глаза.
ресничной мышцы. Ближайшая точка ясного видения находится на расстоянии 10 см от глаза.
Старческая дальнозоркость. Хрусталик с возрастом теряет эластичность, и при изменении натяжения цинковых связок его кривизна меняется мало. Поэтому ближайшая точка ясного видения находится теперь не на расстоянии 10 см от глаза, а отодвигается от него. Близкие предметы при этом видны плохо. Это состояние называется старческой дальнозоркостью, или пресбиопией. Пожилые люди вынуждены пользоваться очками с двояковыпуклыми линзами.
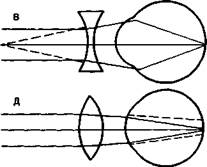
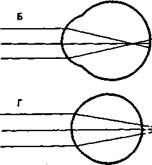
Аномалии рефракции глаза. Две главные аномалии рефракции глаза — близорукость, или миопия, и дальнозоркость, или гиперметропия, — обусловлены не недостаточностью преломляющих сред глаза, а изменением длины глазного яблока (рис. 14.5, А).
Близорукость. Если продольная ось глаза слишком длинная, то лучи от далекого объекта сфокусируются не на сетчатке, а перед ней, в стекловидном теле (рис. 14.5, Б). Такой глаз называется близоруким, или миопическим. Чтобы ясно видеть вдаль, необходимо перед близорукими глазами поместить вогнутые стекла, которые отодвинут сфокусированное изображение на сетчатку (рис. 14.5, В).
Дальнозоркость. Противоположна близорукости дальнозоркость, или гиперметропия. В дальнозорком глазу (рис. 14.5, Г) продольная ось глаза укорочена, и поэтому лучи от далекого объекта фокусируются не на сетчатке, а за ней. Этот недостаток рефракции может быть компенсирован аккомодационным усилием, т. е. увеличением выпуклости хрусталика. Поэтому дальнозоркий человек напрягает аккомодационную мышцу, рассматривая не только близкие, но и далекие объекты. При рассматривании близких объектов аккомодационные усилия дальнозорких людей недостаточны. Поэтому для чтения дальнозоркие люди должны надевать очки с двояковыпуклыми линзами, усиливающими преломление света (рис. 14.5, Д). Гиперметропию не следует путать со старческой дальнозоркостью. Общее у них лишь то, что необходимо пользоваться очками с двояковыпуклыми линзами.
Астигматизм. К аномалиям рефракции относится также астигматизм, т. е. неодинаковое преломление лучей в разных направлениях (например, по горизонтальному и вертикальному меридиану). Астигматизм обусловлен не строго сферической поверхностью роговой оболочки. При астигматизме сильных степеней эта поверхность может приближаться к цилиндрической, что исправляется цилиндрическими очковыми стеклами, компенсирующими недостатки роговицы.
Зрачок и зрачковый рефлекс. Зрачком называют отверстие в центре радужной оболочки, через которое лучи света проходят внутрь глаза. Зрачок повышает четкость изображения на сетчатке, увеличивая глубину резкости глаза. Пропуская только центральные лучи, он улучшает изображение на сетчатке также за счет устранения сферической аберрации. Если прикрыть глаз от света, а затем открыть его, то расширившийся при затемнении зрачок быстро сужается («зрачковый рефлекс»). Мышцы радужной оболочки изменяют величину зрачка, регулируя поток света, попадающий в глаз. Так, на очень ярком свету зрачок имеет минимальный диаметр (1,8 мм), при средней дневной освещенности он расширяется (2,4 мм), а в темноте расширение максимально (7,5 мм). Это приводит к ухудшению качества изображения на сетчатке, но увеличивает чувствительность зрения. Предельное изменение диаметра зрачка изменяет его площадь примерно в 17 раз. Во столько же раз меняется при этом световой поток. Между интенсивностью освещения и диаметром зрачка имеется логарифмическая зависимость. Реакция зрачка на изменение освещенности имеет адаптивный характер, так как в небольшом диапазоне стабилизирует освещенность сетчатки.
В радужной оболочке имеется два вида мышечных волокон, окружающих зрачок: кольцевые (m. sphincter iridis), иннервируемые парасимпатическими волокнами глазодвигательного нерва, а также радиальные (m. dilatator iridis), иннервируемые симпатическими нервами. Сокращение первых вызывает сужение, сокращение вторых — расширение зрачка. Соответственно этому ацетилхолин и эзерин вызывают сужение, а адреналин — расширение зрачка. Зрачки расширяются во время боли, при гипоксии, а также при эмоциях, усиливающих возбуждение симпатической системы (страх, ярость). Расширение зрачков — важный симптом ряда патологических состояний, например болевого шока, гипоксии.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|