- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Рис. 7.2.10. Рефракция глаза и ее аномалии
Рис. 7.2.10. Рефракция глаза и ее аномалии
Приспособление глаза к ясному видению разноудаленных предметов называется аккомодацией. При аккомодации изменяются кривизна хрусталика и его преломляющая способность. В основе аккомодации лежит рефлекторное сокращение цилиарной (ресничной) мышцы, приводящее к изменению выпуклости хрусталика при нечетком изображении рассматриваемого предмета (рис. 7.2.11).
Рефлекторный путь аккомодации начинается от рецепторов сетчатки глаза. Информация от них по зрительным нервам и чувствительным трактам поступает к парасимпатическим ядрам глазодвигательного нерва среднего мозга. От нейронов ядер возбуждение через нервный узел идет к цилиарным мышцам, вызывая их сокращение. Сокращение цилиарных мышц приводит к ослаблению тяги цин- новых связок хрусталика и он вследствие своей эластичности принимает более выпуклую форму. Для нормального глаза дальняя точка ясного видения лежит в бесконечности, поэтому глаз далекие предметы рассматривает без напряжения аккомодации, т.е. без сокращения ресничных мышц. Предметы, расположенные ближе 10 см, видятся человеку с нормальным зрением неясными даже при максимальном сокращении ресничных мышц, т.е. при максимальном аккомодационном усилии.

Рис. 7.2.11. Аккомодация глаза
Зрачковый рефлекс характеризуется изменением диаметра зрачков. Отверстие в центре радужной оболочки — зрачок — пропускает центральные и отклоняет лучи света, поступающие с периферии. Это способствует четкости изображения предмета на сетчатке. Размер зрачка изменяется за счет сокращения мышц радужки. Реакция зрачка на освещенность носит адаптивный характер и регулирует уровень освещенности сетчатки. В темноте диаметр зрачка увеличивается (расширение зрачка), а на свету его диаметр уменьшается (сужение зрачка). Изменения диаметра зрачка происходят рефлек- торно и носят название зрачковый рефлекс (рис. 7.2.12. Зрачковый рефлексе^).
Путь зрачкового рефлекса начинается от рецепторов сетчатки. При увеличении освещенности возбуждение по зрительным нервам и сенсорным трактам поступает к нейронам ядер глазодвигательного нерва, а далее через парасимпатический (цилиарный ганглий) — к радиальным мышцам радужной оболочки. Их сокращение приводит к сужению зрачка.
При уменьшении освещенности возбуждение рецепторов сетчатки передается по зрительным нервам и трактам к нейронам симпатического цилиоспинального центра спинного мозга. Аксоны этих нейронов направляются в верхний шейный узел, а оттуда — к мышцам радужки, расширяющим зрачок. Расширение зрачка может возникать как проявление активации симпатической нервной системы (страх, ярость, боль).
Рецепторный отдел зрительной сенсорной системы. Представлен фоторецепторами сетчатки — палочками и колбочками. В сетчатке глаза человека насчитывается около 6,5 млн колбочек и 120 млн палочек. Каждый фоторецептор состоит из чувствительного к свету наружного сегмента, содержащего зрительный пигмент, и внутреннего сегмента, в состав которого входят органеллы (ядро, митохондрии), участвующие в энергетических процессах фоторецепторной клетки (рис. 7.2.13).
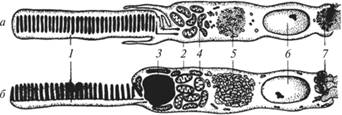
Рис. 7.2.13. Строение палочки (а) и колбочки (6) сетчатки позвоночных:
1,2 — наружный и внутренний сегменты; 3 — масляная капля; 4 — эллипсоид (митохондрии); 5 — параболоид; 6 — ядро; 7 — синаптическая область
В центре пересечения сетчатки с оптической осью глаза находится желтое пятно. Цвет пятна обусловлен высокой концентрацией каротиноидов. В нем имеется углубление — центральная ямка, которая содержит только колбочки. По направлению к периферии сетчатки число колбочек уменьшается, а количество палочек возрастает. Периферические зоны сетчатки содержат исключительно палочки.
Колбочки функционируют в условиях яркой освещенности и воспринимают цвета. Палочки являются рецепторами, которые чувствительны к свету в условиях сумеречного зрения. Место выхода зрительного нерва из глазного яблока не содержит фоторецепторов Ослепое пятно), поэтому оно нечувствительно к свету.
Фоторецепторы сетчатки человека и высших животных располагаются ближе к ее пигментному слою. Он препятствует отражению и рассеиванию светового потока, поглощает продукты деградации фоторецепторов и превращений зрительных пигментов, а также способствует обновлению рецепторного слоя сетчатки. Помимо рецепторов, в сетчатке глаза находятся афферентные нейроны проводникового отдела сенсорной системы, которые в совокупности с фоторецепторными клетками образуют сложную нервную сеть, участвующую в первичной обработке зрительной информации (рис. 7.2.14).
По направлению к стекловидному телу от фоторецепторных клеток расположен слой биполярных нейронов, к которому изнутри примыкают ганглиозные нервные клетки.
Фоторецепторы относятся к вторично-чувствующим, поэтому они образуют синапсы с биполярными клетками. Каждый биполярный нейрон связан со многими палочками и несколькими колбочками, а одна ганглиозная клетка — со многими биполярными
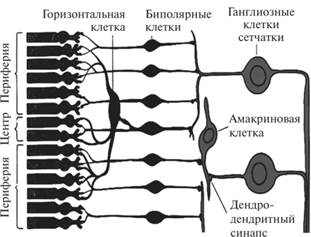
Рис. 7.2.14. Строение сетчатки глаза [4]
клетками. Взаимодействие между соседними биполярными нейронами сетчатки и ганглиозными клетками обеспечивают горизонтальные и амакриновые клетки соответственно.
Фотохимические процессы в сетчатке глаза. Рецепторные клетки сетчатки содержат светочувствительные пигменты (хромопротеиды), которые под влиянием света меняют изомерную форму — обесцвечиваются. В палочках на мембране наружных сегментов содержится родопсин, а в колбочках — иодопсин и другие пигменты. Зрительные пигменты состоят из ретиналя и гликопротеида (опсина). В темноте происходит ресинтез пигментов. При постоянном и равномерном освещении устанавливается динамическое равновесие между скоростью распада и ресинтеза зрительных пигментов. При адаптации к темноте равновесие сдвигается в сторону более высоких концентраций пигмента.
Зрительные пигменты в фоторецепторах связаны с G-белком (трансдуцином). Действие света, например, на родопсин фоторецептора активирует трансдуцин, он активирует фосфодиэстеразу, которая снижает количество цГМФ, поддерживающего Na+-Ka- налы наружной мембраны в открытом состоянии. Натриевая проводимость мембраны фоторецептора снижается, и развивается гиперполяризация фоторецепторной клетки (рецепторный потенциал).
Синаптические окончания фоторецепторных клеток сходятся (конвергируют) на биполярных нейронах сетчатки. Фоторецепторы центральной ямки связаны только с одной клеткой, тогда как в периферических отделах сетчатки происходит конвергенция множества фоторецепторов к одной биполярной клетке, что повышает вероятность обнаружения первичного сигнала.
При действии света на сетчатку в биполярных клетках, также, как и в рецепторах, возникает локальное возбуждение, передающееся затем на ганглиозные клетки (рис. 7.2.15. Электрическая активность различных клеток сетчатки под действием светаС^У).
В одних биполярных клетках в ответ на включение и выключение света возникает медленная длительная деполяризация, в других включение света вызывает гиперполяризацию, а выключение — деполяризацию. Это связано с наличием в биполярных клетках разных типов постсинаптических рецепторов: возбуждающих или тормозных.
Проводниковый отдел зрительной сенсорной системы. Первыми сенсорными нейронами проводникового отдела зрительной системы являются ганглиозные клетки сетчатки. К ним подходят отростки биполярных клеток. Число отростков биполярных клеток, контактирующих с ганглиозными клетками, неодинаково: чем ближе ганглиозные клетки к центральной ямке, тем меньше биполярных клеток контактирует с одной клеткой.
В желтом пятне число колбочек практически равно числу биполярных и ганглиозных клеток. С этим связывают высокую остроту зрения в центральных отделах сетчатки. На периферии сетчатки к одной ганглиозной клетке через биполярные клетки одномоментно поступают сигналы почти от 600 палочек. В результате суммации импульсов от множества палочек происходит более интенсивная стимуляция ганглиозных клеток, что повышает чувствительность периферических зон сетчатки к свету. Генерация импульсов в ганглиозных клетках происходит даже в полной темноте.
В ответ на световой стимул ганглиозные клетки изменяют частоту генерации потенциалов действия. Реакция ганглиозной клетки зависит от освещенности ее рецептивного поля (рис. 7.2.16. Ответы ганглиозных клеток на раздражение рецепторных полей с оп-цен- тром и off-центромС^).
Ганглиозные клетки реагируют на освещение центра их рецептивного поля двояко: возбуждением (клетки с оп-центром) или торможением (клетки с off-центром). Горизонтальные и амакриновые клетки осуществляют латеральное торможение между биполярными или ганглиозными клетками, регулируя величину рецептивных полей проводниковых нейронов.
Аксоны ганглиозных клеток формируют зрительный нерв. По нему потенциалы действия от ганглиозных клеток поступают в головной мозг. В головном мозге волокна зрительного нерва образуют неполный перекрест, поэтому сетчатка одного глаза имеет проекцию на противоположной и той же сторонах в вышележащих структурах мозга (рис. 7.2.17).
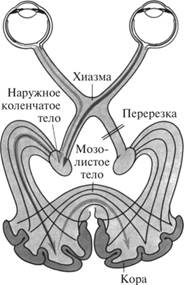
Рис. 7.2.17. Специфический зрительный путь и корковые проекции
Большая часть волокон в составе зрительных трактов поступает в подкорковые центры зрительной сенсорной системы, а оттуда в первичную проекционную зону затылочной области коры. Часть волокон направляется в ядра среднего мозга, в которых сосредоточены нейроны, ответственные за движение глазных яблок и изменение диаметра зрачка.
Корковый отдел зрительной сенсорной системы. Локализуется в затылочной доле коры большого мозга (рис. 7.2.18. Корковый отдел зрительной сенсорной системьк?2)). В зрительной коре существуют функционально различные группы клеток: простые, сложные и сверхсложные. Простая клетка реагирует только на полоску света определенной ориентации в строго заданном месте ее рецептивного поля. Сложные нейроны реагируют на строго ориентированные стимулы, которые могут быть локализованы в разных областях рецептивного поля. Они участвуют в определении угла наклона или направления движения линии в поле зрения. Сверхсложные нейроны зрительной коры включаются при детектировании сложных признаков раздражителя, например пересекающихся линий и углов.
Цветовое зрение. В восприятии цвета участвуют первичные и вторичные фоторецепторные механизмы. С помощью первичных механизмов происходит оценка спектральных характеристик света. Различение цветов осуществляют воспринимающие цвет фоторецепторы. Они избирательно реагируют на разные участки спектра. Вторичные (нервные) механизмы перекодируют первичную информацию о цвете, поступающую от фоторецепторов.
Цветовое зрение связано с наличием в сетчатке глаза фоторецепторов трех видов — колбочек, раздельно воспринимающих красный, зеленый и сине-фиолетовый цвета. Каждый цвет воздействует на колбочки всех трех типов. Однако в зависимости от спектральных характеристик светового раздражителя интенсивность их возбуждения неодинаковая. Комбинации возбуждения различных колбочек суммируются зрительными нейронами, и в коре головного мозга формируют ощущение цветов и оттенков. Равномерное возбуждение всех трех типов рецепторов дает ощущение белого света.
Правильное восприятие цвета имеет большое значение в повседневной жизни людей, а также для лиц, которые по роду своей профессии контактируют с цветовыми индикаторами (водители, машинисты, авиадиспетчеры, летчики и др.). Исследование цветового зрения проводят с помощью полихроматических таблиц Е.Б. Раб- кина.
Острота зрения. Под остротой зрения понимают способность глаза воспринимать две светящиеся точки раздельно при минимальном расстоянии между ними. Нормальный глаз различает две точки раздельно под углом зрения в одну минуту. Для раздельного видения двух точек необходимо, чтобы между возбужденными колбочками находилась минимум одна невозбужденная колбочка. Диаметр колбочки равен 3 мкм. Для раздельного видения двух точек необходимо, чтобы расстояние между изображениями этих точек на сетчатке составляло не менее 4 мкм, т.е. величины изображения при угле зрения в одну минуту. Если угол зрения будет менее одной минуты, то две светящиеся точки сливаются в одну. Определение остроты зрения осуществляется с помощью буквенных или различного вида фигурных стандартных таблиц. Острота зрения выражается в относительных величинах.
Поле зрения. Полем зрения называется пространство, видимое глазом при фиксации взгляда в одной точке. Если фиксировать взгляд на каком-либо предмете, то изображение попадает в область желтого пятна — зону наилучшего видения. Предметы, изображения которых проецируются на остальные места сетчатки, воспринимаются периферическим зрением. Различают цветовое (хроматическое) и бесцветное (ахроматическое) поля зрения. Ахроматическое поле зрения больше хроматического, так как оно обусловлено деятельностью палочек, расположенных преимущественно на периферии сетчатки. Для различных цветов поле зрения неодинаково, больше всех оно для желтого цвета, а самое узкое — для зеленого. Определяется поле зрения с помощью периметра.
Восприятие пространства и величины предмета. Восприятие глубины пространства и оценка расстояния до объекта возможны как при зрении одним глазом (монокулярное зрение), так и двумя глазами (бинокулярное зрение). При бинокулярном зрении оценка расстояния гораздо точнее. В этом случае изображения предмета имеются на сетчатках обоих глаз, но у человека не возникает ощущения двух предметов, так как световые проекции от предмета попадают на соответственные, или идентичные, участки сетчатки, и в восприятии человека эти два изображения сливаются в одно. В этом легко убедиться, если надавить слегка на один глаз сбоку: изображение раздваивается, так как смещается проекция предмета на сетчатке. Если рассматривать разноудаленные предметы двумя глазами, их изображения попадают на неидентичные (диспаратные) участки сетчатки, смещенные относительно друг друга. Анализ этого смещения в зрительной коре головного мозга обеспечивает восприятие глубины пространства.
При монокулярном зрении определенное значение в оценке близких расстояний имеет явление аккомодации. Чтобы оценить расстояние до предмета, важно также увеличение размера его проекции на сетчатке при приближении предмета к глазу. Размер предмета оценивается как функция двух переменных: размера изображения на сетчатке; расстояния предмета от глаза. Если расстояние до незнакомого предмета вследствие недостаточной его рельефности оценить трудно, то возможны ошибки и в определении размера предмета.
Слуховая сенсорная система. Слух позволяет человеку ориентироваться во всем многообразии звуковых раздражителей внешней среды. Слух тесно связан с речевой функцией. Он необходим для голосовой связи и умения говорить.
Вестибулярная сенсорная система обеспечивает ориентацию человека в пространстве. Она формирует ощущения, которые возникают при прямолинейном и вращательном движениях, а также при изменениях положения головы. Орган слуха и равновесия анатомически и функционально тесно взаимосвязаны. Орган слуха включает в себя наружное, среднее и часть внутреннего уха — улитку, которая представлена улитковым лабиринтом. Орган равновесия расположен только во внутреннем ухе и включает такие части лабиринта, как преддверие и полукружные каналы (см. 7.2.19).
Слуховая сенсорная система — это второй по информационной значимости канал связи организма с внешней средой.

Рис. 7.2.19. Строение органа слуха и равновесия (фронтальный распил):
· 7 — молоточек; 2 — полукружные каналы; 3 — преддверие; 4 — улитка; 5 — преддверно-улитковый нерв; 6 — внутренняя сонная артерия; 7 — слуховая труба; 8 — барабанная полость; 9 — барабанная перепонка; 10 — наружный слуховой проход;
· 7 7 — ушная раковина [6]
Строение органа слуха. Наружное ухо состоит из ушной раковины и наружного слухового прохода. Ушная раковина построена из эластического хряща и имеет форму воронки. Она «собирает» звуковые колебания и направляет их в наружный слуховой проход. Наружный слуховой проход по форме напоминает узкую, слепо заканчивающуюся трубку длиной 20—25 мм. Поверхность наружного слухового прохода выстлана кожей, в которой находятся серные железы. Слуховой проход служит звуковым резонатором. Наружное ухо обеспечивает лучшее восприятие звуков, идущих в одном направлении.
Наружное и среднее ухо разделяет барабанная перепонка. Барабанная перепонка — плотная мембрана, сплетенная из соединительнотканных волокон. Волокна мембраны не имеют общего направления и натянуты неравномерно. Благодаря такому строению барабанная перепонка не обладает собственным периодом колебаний, наличие которого приводило бы к искажению воспринимаемого звука. Колебания барабанной перепонки возникают под действием звуковых волн, проходящих через наружный слуховой проход.
Среднее ухо расположено внутри височной кости. Оно представлено барабанной полостью (до 1 см3), слуховой трубой и ячейками костного отростка. Барабанная полость выстлана слизистой оболочкой и содержит три слуховые косточки и две мышцы. Слуховые косточки — молоточек, наковальня и стремечко — последовательно, подвижно (суставами) соединены между собой. Молоточек с помощью своей рукоятки неподвижно связан с барабанной перепонкой, а основание стремечка закрывает овальное окно в преддверии костного лабиринта. Одна из мышц осуществляет напряжение барабанной перепонки, вторая — обеспечивает движение стремечка в овальном окне. Барабанная полость соединена с носоглоткой слуховой (евстахиевой) трубой. Посредством евстахиевой трубы происходит выравнивание давления в барабанной полости с атмосферным давлением, что создает благоприятные условия для колебаний барабанной перепонки.
Звуковые колебания от барабанной перепонки во внутреннее ухо передаются по слуховым косточкам. Они представляют собой косточковую рычажную систему, которая дает 22-кратное увеличение общего давления на жидкость улитки по сравнению с силой давления звуковых волн на барабанную перепонку. Изменения давления и колебания столба жидкости при этом в улитке возможны благодаря наличию в стенке, отделяющей среднее ухо от внутреннего, помимо овального, круглого окна, закрытого эластичной мембраной.
При отсутствии барабанной перепонки и слуховых косточек улитка сохраняет способность воспринимать прямые звуковые сигналы. Слуховая чувствительность уха при этом резко снижается — умеренно громкий звук воспринимается как едва слышимый. Передача звуковых колебаний возможна и через кости черепа. При закрытом слуховом проходе звук будет слышен, если ножку звучащего камертона приложить к темени или сосцевидному отростку.
Внутреннее ухо и восприятие звуков. Внутренне ухо представляет собой сложную по форме систему каналов, называемую лабиринтом, которые заполнены специальной жидкостью. Различают костный лабиринт, внутри которого находится перепончатый лабиринт. Пространство между ними заполнено перилимфой, а внутри перепончатого лабиринта содержится эндолимфа. В костном лабиринте различаю три части: улитку, преддверие и костные полукружные каналы. Улитка относится к органу слуха, а преддверие и костные полукружные каналы — к органу равновесия.
Улитка — спирально закрученный костный канал, имеющий 2,5 завитка. Канал улитки разделен на три хода: верхний, средний и нижний (рис. 7.2.20).
Верхний и нижний ходы сообщаются между собой через отверстие (геликотрему), образуя заполненное перилимфой пространство, которое ограничено мембранами овального и круглого окон. Между этими ходами расположен средний (перепончатый) канал. Он заполнен эндолимфой и с соседними ходами улитки не сообщается. Внутри среднего канала на основной мембране расположен звуковоспринимающий аппарат — спиральный (кортиев) орган, который содержит рецепторные волосковые клетки (рис. 7.2.21).
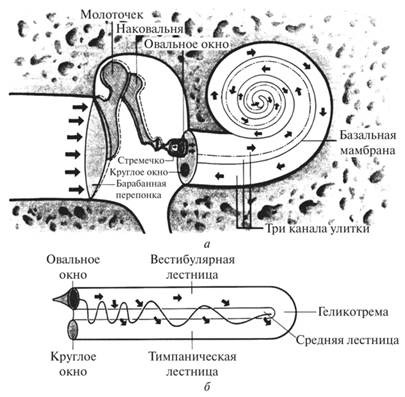
Рис. 7.2.20. Каналы улитки:
а — среднее и внутреннее ухо в разрезе (по П. Линдсею и Д. Норманну, 1974); б — распространение звуковых колебаний в улитке

Рис. 7.2.21. Кортиев орган
Рецепторные волосковые клетки трансформируют механические колебания в электрические потенциалы, что приводит к возбуждению волокон слухового нерва. Один полюс волосковой клетки фиксирован на основной мембране, а второй находится в полости перепончатого (среднего) канала. При действии звуков основная мембрана колеблется, волоски рецепторных клеток деформируются, что сопровождается деполяризацией мембраны — генерацией рецепторного потенциала и возбуждением волокон слухового нерва. Передача возбуждения с волосковой клетки на первый нейрон проводниковой системы происходит через синапсы.
Первый нейрон проводникового отдела слуховой сенсорной системы представлен биполярной клеткой, расположенной в спиральном ганглии улитки. Аксоны биполярных клеток образуют слуховой нерв и заканчиваются на нейронах кохлеарных ядер продолговатого мозга (второй нейрон слухового пути). Отсюда возбуждение поступает в верхние оливы, в задние бугры четверохолмия и затем — в медиальные коленчатые тела таламуса (рис. 7.2.22. Схема связей в слуховой сенсорной системе
Аксоны таламических нейронов проводят возбуждение в корковый отдел, расположенный в верхней части височной доли. Центры первичной слуховой коры формируют способность человека локализовать звук и определять направление его смещения (пространственный слух), а также интерпретировать простые звуковые образы. Ассоциативные зоны слуховой коры ответственны за распознавание значений слышимых слов.
Звуковые ощущения. Человек может воспринимать звуки с частотой колебаний от 16 до 20 кГц. Минимальная сила звука, слышимого человеком, называется абсолютной слуховой чувствительностью. Пороги слышимости изменяются в зависимости от частоты звука. Максимальной чувствительностью слух людей обладает в области диапазона частот человеческого голоса — от 1 до 4 кГц.
Восприятие частоты звуковых колебаний происходит с помощью нескольких механизмов. Звук вызывает колебания основной мембраны улитки, распространяющиеся от ее основания к вершине. Звуковая волна имеет наибольшую амплитуду колебаний на строго определенном участке мембраны в зависимости от ее частоты. Высокочастотные колебания вызывают колебания мембраны у основания улитки, а низкочастотные волны — ближе к вершине улитки. Звуки низкой частоты воспринимаются рецепторами, расположенными ближе к верхушке улитки, а звуки высокой частоты — рецепторами, расположенными ближе к основанию улитки. От каждого типа рецепторов возбуждение проводится по слуховым проводящим путям в нейроны определенных колонок слуховой коры головного мозга, ответственных за восприятие звука конкретной частоты. Тон звука также кодируется частотой потенциалов действия, распространяющихся по слуховому нерву.
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|