- Автоматизация
- Антропология
- Археология
- Архитектура
- Биология
- Ботаника
- Бухгалтерия
- Военная наука
- Генетика
- География
- Геология
- Демография
- Деревообработка
- Журналистика
- Зоология
- Изобретательство
- Информатика
- Искусство
- История
- Кинематография
- Компьютеризация
- Косметика
- Кулинария
- Культура
- Лексикология
- Лингвистика
- Литература
- Логика
- Маркетинг
- Математика
- Материаловедение
- Медицина
- Менеджмент
- Металлургия
- Метрология
- Механика
- Музыка
- Науковедение
- Образование
- Охрана Труда
- Педагогика
- Полиграфия
- Политология
- Право
- Предпринимательство
- Приборостроение
- Программирование
- Производство
- Промышленность
- Психология
- Радиосвязь
- Религия
- Риторика
- Социология
- Спорт
- Стандартизация
- Статистика
- Строительство
- Технологии
- Торговля
- Транспорт
- Фармакология
- Физика
- Физиология
- Философия
- Финансы
- Химия
- Хозяйство
- Черчение
- Экология
- Экономика
- Электроника
- Электротехника
- Энергетика
Комментарии 3 страница
Митоз — один из фундаментальных процессов онтогенеза (жизни индивидуального организма). Митотическое деление обеспечивает рост многоклеточных эукариот за счёт увеличения популяций клеток тканей. У растений в результате митотического деления клеток образовательных тканей (меристем) увеличивается количество клеток тканей. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений[2].
На основании морфологических особенностей митоз условно подразделяется на стадии: профазу, прометафазу, метафазу, анафазу, телофазу.
Продолжительность митоза в среднем составляет 1−2 часа[1][3]. Митоз клеток животных, как правило, длится 30−60 минут, а растений — 2−3 часа[4]. За 70 лет в теле человека суммарно осуществляется порядка 1014 клеточных делений[5].
Митоз происходит только в клетках эукариот (ядерных). Клетки прокариот (безъядерных) делятся другим, бинарным, способом. Митоз отличается для разных организмов[6]. Так, например, процесс для клеток животных является «открытым», а для клеток грибов — «закрытым» (при котором хромосомы делятся в целом клеточном ядре)[7]. У человека все клетки, кроме гамет, производятся митозом. Гаметы производятся мейозом.
История исследования[править | править код]
Первые описания фаз митоза и установление их последовательности были предприняты в 1870—1880-х годах. В конце 1870-х — начале 1880-х годов немецкий гистолог Вальтер Флемминг для обозначения процесса непрямого деления клетки ввёл термин «митоз»[8].
Первые неполные описания, касающиеся поведения и изменения ядер в делящихся клетках, встречаются в работах учёных начала 1870-х годов. В работе русского ботаника Эдмунда Руссова, датируемой 1872 годом, отчётливо описаны и изображены метафазные и анафазные пластинки, состоящие из отдельных хромосом[9]. Годом позже немецкий зоолог Антон Шнейдер ещё более отчётливо и последовательно, но, конечно, не совсем полно описал митотическое деление на примере дробящихся яиц Mesostoma ehrenbergii (нид.)русск.[10]. В его работе, в сущности, описаны и проиллюстрированы в правильной последовательности основные фазы митоза: профаза, метафаза, анафаза (ранняя и поздняя). В 1874 году московский ботаник И. Д. Чистяков также наблюдал отдельные фазы клеточного деления в спорах плаунов и хвощей. Несмотря на первые успехи ни Руссову, ни Шнейдеру, ни Чистякову не удалось дать чёткое и последовательное описание митотического деления[11].
В 1875 году вышли работы, содержащие более детальные описания митозов. Отто Бючли дал описание цитологических картин в дробящихся яйцах круглых червей и моллюсков и в сперматогенных клетках насекомых. Эдуард Страсбургер исследовал митотическое деление в клетках зелёной водоросли спирогиры, в материнских клетках пыльцы лука и в материнских споровых клетках плауна. Ссылаясь на работу Отто Бючли и основываясь на собственных исследованиях, Эдуард Страсбургер обратил внимание на единство процессов клеточного деления в растительных и животных клетках[12].
К концу 1878 — началу 1879 года появились подробные работы В. Шлейхера (о делении хрящевых клеток амфибий), В. Флемминга (о размножении клеток в разных тканях саламандры и её личинок), П. И. Перемежко (о делении клеток в эпидермисе личинок тритона). В своей работе в 1879 году Шлейхер предложил термин «кариокинез» для обозначения сложных процессов клеточного деления, подразумевая перемещения составных частей ядра[13]. Вальтер Флемминг впервые для обозначения непрямого деления клетки ввёл термин «митоз», который впоследствии стал общепринятым[8]. Также Флеммингу принадлежит окончательная формулировка определения митоза как циклического процесса, завершающегося разделением хромосом между дочерними клетками[14].
· 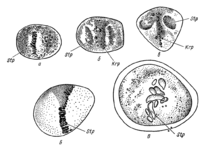
Деление клеток по Э. Руссову (1872)
· 
Деление клеток по Э. Страсбургеру (1875)
· 
Деление клеток по В. Флеммингу (1882)
· 
Деление клеток по Э. Б. Уилсону (1900)
В 1880 году О. В. Баранецкий установил спиральное строение хромосом. В ходе дальнейших исследований были развиты представления о спирализации и деспирализации хромосом во время митотического цикла[14]. В начале 1900-х годов хромосомы были идентифицированы в качестве носителей наследственной информации, что в дальнейшем дало объяснение биологической роли митоза, заключающейся в образовании генетически идентичных дочерних клеток.
В 1970-х годах началась расшифровка и детальное изучение регуляторов митотического деления[15], благодаря серии экспериментов по слиянию клеток, находящихся на разных этапах клеточного цикла. В тех опытах, когда клетку в М-фазе объединяли с клеткой, находящейся в любой из стадий интерфазы (G1, S или G2), интерфазные клетки переходили в митотическое состояние (начиналась конденсация хромосом и распадалась ядерная мембрана)[16]. В итоге был сделан вывод, что в цитоплазме митотической клетки присутствует фактор (или факторы), стимулирующий митоз[17], или, иначе, М-стимулирующий фактор (англ.)русск. (МСФ, от англ. M-phase-promoting factor, MPF)[18].
Впервые «фактор стимуляции митоза» был открыт в зрелых неоплодотворенных яйцах шпорцевой лягушки, находящихся в М-фазе клеточного цикла. Цитоплазма такого яйца, инъецированная в ооцит, приводила к преждевременному переходу в М-фазу и к началу созревания ооцита (первоначально сокращение MPF означало Maturation promoting factor (англ.)русск., что переводится как «фактор, способствующий созреванию»). В ходе дальнейших экспериментов были установлены универсальное значение и вместе с тем высокая степень консервативности «фактора стимуляции митоза»: экстракты, приготовленные из митотических клеток весьма разнообразных организмов (млекопитающих, морских ежей, моллюсков, дрожжей), при введении в ооциты шпорцевой лягушки переводили их в М-фазу[19].
В ходе последующих исследований выяснилось, что фактор, стимулирующий митоз, представляет собой гетеродимерный комплекс, состоящий из белка циклина и зависимой от циклина протеинкиназы. Циклин является регуляторным белком и обнаруживается у всех эукариот. Его концентрация периодически возрастает в течение клеточного цикла, достигая максимума в метафазе митоза. С началом анафазы наблюдается резкое сокращение концентрации циклина, вследствие его расщепления с помощью сложных белковых протеолитических комплексов — протеасом. Зависимая от циклина протеинкиназа представляет собой фермент (фосфорилазу), модифицирующий белки за счёт переноса фосфатной группы от АТФ на аминокислоты серин и треонин. Таким образом с установления роли и структуры основного регулятора митотического деления начались исследования тонких регуляторных механизмов митоза, которые продолжаются до настоящего времени.
Аппарат клеточного деления[править | править код]
Деление всех эукариотических клеток сопряжено с формированием специального аппарата клеточного деления. Активная роль в митотическом делении клеток зачастую отведена цитоскелетным структурам. Универсальным как для животных, так и для растительных клеток является двухполюсное митотическое веретено, состоящее из микротрубочек и связанных с ними белков[20]. Веретено деления обеспечивает строго одинаковое распределение хромосом между полюсами деления, в области которых в телофазе образуются ядра дочерних клеток.
Ещё одна не менее важная структура цитоскелета отвечает за разделение цитоплазмы (цитокинез) и, как следствие, за распределение клеточных органелл. В животных клетках за цитокинез отвечает сократимое кольцо из актиновых и миозиновых филаментов. В большинстве клеток высших растений из-за наличия жёсткой клеточной стенки цитокинез протекает с образованием клеточной пластинки в плоскости между двумя дочерними клетками. При этом область образования новой клеточной перегородки определяется заранее предпрофазным пояском из актиновых микрофиламентов, а поскольку актин участвует также в формировании клеточных септ у грибов, возможно, что он направляет цитокинез у всех эукариот[21].
Веретено деления[править | править код]
Основная статья: Веретено деления
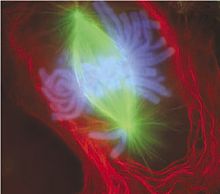
Поздняя метафаза митоза в клетке лёгкого тритона (использованы иммунофлуоресцентные красители)[22]. Чётко просматривается веретено деления, образованное микротрубочками (зелёные), и хромосомы (синие)
Формирование веретена деления начинается в профазе. В его образовании принимают участие полярные тельца (полюса) веретена и кинетохоры хромосом, и те и другие взаимодействуют с микротрубочками — биополимерами, состоящими из субъединиц тубулина. Главным центром организации микротрубочек (ЦОМТ) во многих эукариотических клетках является центросома — скопление аморфного фибриллярного материала, причём в большинстве животных клеток в состав центросом также входят пары центриолей[23]. Во время интерфазы ЦОМТ, как правило, располагающийся вблизи клеточного ядра, инициирует рост микротрубочек, расходящихся к периметру клетки и образующих цитоскелет. В S-фазе материал центросомы удваивается, а в профазе митоза начинается расхождение дочерних центросом. От них в свою очередь «отрастают» микротрубочки, которые удлиняются вплоть до соприкосновения друг с другом, после чего центросомы расходятся. Затем, в прометафазе, после разрушения ядерной мембраны, микротрубочки проникают в область клеточного ядра и взаимодействуют с хромосомами. Две дочерние центросомы теперь называют полюсами веретена[24].
По морфологии различают два типа митотического веретена: астральный (или конвергентный) и анастральный (дивергентный)[~ 1][26].
Астральный тип митотической фигуры, характерный для животных клеток, отличают благодаря небольшим зонам, на полюсах веретена, в которых сходятся (конвергируют) микротрубочки. Зачастую центросомы, располагающиеся в области полюсов астрального веретена, содержат центриоли. От полюсов деления также расходятся во всех направлениях радиальные микротрубочки, не входящие в состав веретена, а образующие звездчатые зоны — цитастеры.
Анастральный тип митотической фигуры отличается широкими полярными областями веретена, так называемыми полярными шапочками, в их состав не входят центриоли. Микротрубочки при этом расходятся широким фронтом (дивергируют) от всей зоны полярных шапочек. Этот тип митотической фигуры также отличает отсутствие цитастеров. Анастральный тип митотического веретена наиболее характерен для делящихся клеток высших растений, хотя иногда наблюдается и в некоторых клетках животных.
Микротрубочки[править | править код]
Основная статья: Микротрубочки
Микротрубочки — динамичные структуры, принимающие активное участие в построении веретена деления во время митоза. Химически они представляют собой биополимеры, состоящие из субъединиц белка тубулина. Количество микротрубочек в клетках различных организмов может значительно отличаться. В метафазе веретено деления в клетках высших животных и растений может содержать до нескольких тысяч микротрубочек, тогда как у некоторых грибов их всего около 40[24].
Митотические микротрубочки веретена деления «динамически нестабильны». Их «положительные» или «плюс-концы», расходящиеся во всех направлениях от центросом резко переходят от равномерного роста к стремительному укорочению, при котором часто деполимеризуется вся микротрубочка. Согласно этим данным, образование митотического веретена объясняется селективной (выборочной) стабилизацией микротрубочек, взаимодействующих в экваториальной области клетки с кинетохорами хромосом и с микротрубочками, идущими от противоположного полюса деления. Данная модель объясняет характерную двухполюсную фигуру митотического веретена[24].
Центромеры и кинетохоры[править | править код]
Основные статьи: Центромера и Кинетохор
Центромеры — специализированные последовательности ДНК, необходимые для связывания с микротрубочками веретена деления и для последующего расхождения хромосом. В зависимости от локализации различают несколько типов центромер. Для голоцентрических центромер характерно образование связей с микротрубочками веретена по всей длине хромосомы (некоторые насекомые, нематоды, некоторые растения). В противоположность голоцентрическим моноцентрические центромеры служат для связи с микротрубочками в единственной области хромосомы[26].
В центромерной области обычно располагаются кинетохоры хромосом — сложные белковые комплексы, морфологически очень сходные по своей структуре для различных групп эукариот, как, например, для диатомовых водорослей, так и для человека[27]. Обычно на каждую хроматиду (хромосому) приходится по одному кинетохору. На электронных микрофотографиях кинетохор обычно выглядит как пластинчатая трёхслойная структура[28]. Порядок слоев следующий: внутренний плотный слой, примыкающий к телу хромосомы; средний рыхлый слой; внешний плотный слой, от которого отходит множество фибрилл, образуя т. н. фиброзную корону кинетохора.
К основным функциям кинетохора относят: закрепление микротрубочек веретена деления, обеспечение движения хромосом во время митоза при участии микротрубочек, связывание между собой сестринских хроматид и регуляцию их последующего разделения в анафазе митоза[29]. Минимально достаточно одной микротрубочки (например, для дрожжей) ассоциированной с кинетохором, чтобы обеспечить движение хромосомы. Однако с одним кинетохором могут быть связаны целые пучки, состоящие из 20—40 микротрубочек (например, у высших растений или человека), чтобы обеспечить расхождение хромосом к полюсам клетки[28][29].
Продолжительность митоза[править | править код]
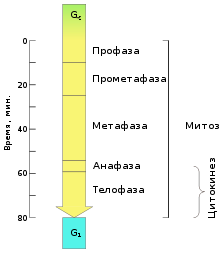
Временной ход митоза и цитокинеза, типичный для клетки млекопитающего. Точные числа для разных клеток различны. Цитокинез берёт своё начало в анафазе и завершается, как правило, к окончанию телофазы
Собственно митоз зачастую протекает сравнительно быстро. Средняя продолжительность составляет 1—2 часа,[1][3] что занимает всего около 10 % времени клеточного цикла. К примеру, у делящихся клеток меристемы корней интерфаза составляет 16—30 часов, а митоз длится всего 1—3 часа. Для эпителиальных клеток кишечника мыши интерфазный период составляет порядка 20—22 часов, а митоз продолжается в течение 1 часа.[30] В клетках животных митоз обычно протекает быстрее и длится в среднем 30—60 минут, в то время как в растительных клетках средняя продолжительность митоза составляет 2—3 часа.[4] Известны исключения с противоположными показателями. К примеру, в животных клетках продолжительность митоза может достигать 3,8 часов (эпидермис мыши). Или же встречаются растительные объекты с длительностью митоза в 5 минут (Chilomonas).[31] Наиболее интенсивно митоз протекает в эмбриональных клетках (10—40 минут в дробящихся яйцеклетках).
Длительность митоза находится в зависимости от целого ряда факторов: размеров делящейся клетки, её плоидности, числа ядер. Частота клеточных делений также зависит от степени дифференцировки клеток и специфики выполняемых функций. Так, нейроны или клетки скелетной мышцы человека не делятся совсем; клетки печени обычно делятся раз в один или два года, а некоторые эпителиальные клетки кишечника делятся чаще, чем 2 раза в сутки.[32]
Темп клеточного деления зависит также от условий окружающей среды, в частности, от температуры. Повышение температуры окружающей среды в физиологических пределах повышает скорость митоза, что может быть объяснено обычной закономерностью кинетики химических реакций.[33]
Фазы митоза[править | править код]
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой (от слова «митоз»). М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую.[23][30] Первые пять — профаза, прометафаза (метакинез), метафаза, анафаза и телофаза (или цитотомия) — составляют митоз,[~ 2] а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы.
Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны стадии, сопряженные с процессами внутриклеточного синтеза: профаза (2—270 минут) и телофаза (1,5—140 минут). Наиболее быстротечны фазы митоза, в ходе которых происходит движение хромосом: метафаза (0,3—175 минут) и анафаза (0,3—122 минуты). Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут.[35]
Препрофаза[править | править код]

Деление растительной клетки с крупной центральной вакуолью
1) Клетка с крупной центральной вакуолью и ядром, оттеснённым на периферию клетки.
2) Полоски цитоплазмы рассекают вакуоль, обеспечивая перемещение ядра к центру клетки.
3) и 4) Полоски цитоплазмы объединяются в плоскости деления клетки с образованием фрагмосомы.
5) На этапе цитокинеза в области фрагмосомы образуется фрагмопласт и новая клеточная стенка
Основная статья: Препрофаза
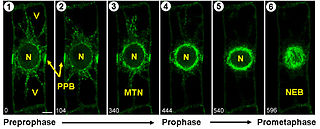
Начальные стадии митоза растительной клетки:
1) и 2) Оформление препрофазного кольца.
3) Нуклеация микротрубочек вокруг ядра.
4) Препрофазное кольцо начинает исчезать.
5) Микротрубочки вокруг ядра образуют «профазное веретено».
6) Ядерная мембрана распадается и микротрубочки веретена направляются к хромосомам.
Условные обозначения: N — ядро; V — вакуоль; PPB — препрофазное кольцо; MTN — начало скопления микротрубочек; NEB — распад ядерной оболочки; микротрубочки окрашены зелёным цветом
Препрофаза — редко используемый термин[36] для обозначения дополнительной стадии митоза растительной клетки. К основным событиям препрофазы можно отнести: оформление препрофазного кольца, образование фрагмосомы и начало нуклеации микротрубочек вокруг клеточного ядра. Несмотря на наличие термина «препрофаза», перечисленные события чаще рассматриваются в составе фазы G2[36][37][38] или в составе профазы.[36][39]
В клетках, богатых вакуолями, в ходе препрофазы образуется фрагмосома — одна из структур, определяющих плоскость деления растительной клетки. Фрагмосома представляет собой слой цитоплазмы, пересекающий вакуоль в плоскости клеточного деления.[40] Ядро в клетках с крупной центральной вакуолью обычно располагается на периферии. Во время препрофазы оно перемещается в область фрагмосомы. В ходе перемещения ядра вакуоль рассекается полосками цитоплазмы, содержащими элементы цитоскелета. Во фрагмосоме также образуется митотическое веретено. Во время цитокинеза в области фрагмосомы формируется фрагмопласт и новая клеточная стенка.
Одновременно с фрагмосомой образуется препрофазное кольцо, и обе структуры располагаются в одной плоскости.[41] Препрофазное кольцо — это кольцеобразное скопление микротрубочек и актиновых филаментов вблизи клеточной мембраны в плоскости деления растительной клетки. Ядро располагается в центре препрофазного кольца и связано с ним радиально расходящимися микротрубочками. Внешне данная структура напоминает колесо с ободом и спицами из микротрубочек и актиновых филаментов, а также с ядром на месте втулки.[41] Структура кольца также обогащена элементами ЭПР и везикулами аппарата Гольджи.
Препрофазное кольцо формируется перед профазой митоза. После начала профазы микротрубочки кольца деполимеризуются и далее участвуют в образовании веретена деления. Функции препрофазного кольца пока не ясны. Однако подмечено, что цитокинез растительной клетки происходит в плоскости определённой положением препрофазного кольца.[36] При симметричном делении кольцо формируется в середине, а при асимметрическом делении образуется ближе к одному концу клетки.[41]
Профаза[править | править код]
Основная статья: Профаза
К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки.[42] Распад ядрышка в профазе является характерной, но необязательной для всех клеток особенностью.[43]
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры, необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.[44]

Профаза
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено — одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца (центросомы), микротрубочки и кинетохоры хромосом.[26]
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут (в интерфазе) до 15 секунд.[24][44] Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками.[44] Полимеризующиеся плюс-концы («+»-концы) являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка.[24] Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.[~ 3]
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами («−»-концами) микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.[46]
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.[47]
Прометафаза[править | править код]
Основная статья: Прометафаза

Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны.[42] Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают.[48] После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин.[48] Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом.[48][49] В результате такого взаимодействия кинетохорные микротрубочки (связанные с кинетохором) стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена.[45] Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.[50]
В клетках млекопитающих прометафаза протекает, как правило, в течение 10—20 минут.[49] В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона — около 30 минут.[51] В дрожжевых клетках невозможно чётко разграничить стадии профазы и прометафазы по причине сохранения ядерной оболочки в процессе деления. Аналогичным образом, частичное или более позднее разрушение ядерной мембраны затрудняет разграничение стадий профазы и прометафазы в клетках Drosophila и C. elegans. В подобных случаях для описания всех ранних событий митотического деления используется обобщающий термин «профаза».[42]
Метафаза[править | править код]
Основная статья: Метафаза

Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена (а не всей клетки[52]) примерно на равном расстоянии от обоих полюсов деления, образуя метафазную (экваториальную) пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи — к периферии клетки (фигура «материнской звезды»). В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.[53][54] В дрожжевых клетках хромосомы тоже не выстраиваются в экваториальной плоскости, а располагаются произвольно вдоль волокон веретена деления.[42]
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.[55]
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума.[53]
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.[53]
Анафаза[править | править код]
Основная статья: Анафаза
|
|
|
© helpiks.su При использовании или копировании материалов прямая ссылка на сайт обязательна.
|